

Combined laryngeal inflammation and trauma mediate long-lasting immunoreactivity response in the brainstem sensory nuclei in the rat
- PMID: 23162441
- PMCID: PMC3498623
- DOI: 10.3389/fnint.2012.00097
Combined laryngeal inflammation and trauma mediate long-lasting immunoreactivity response in the brainstem sensory nuclei in the rat
Abstract
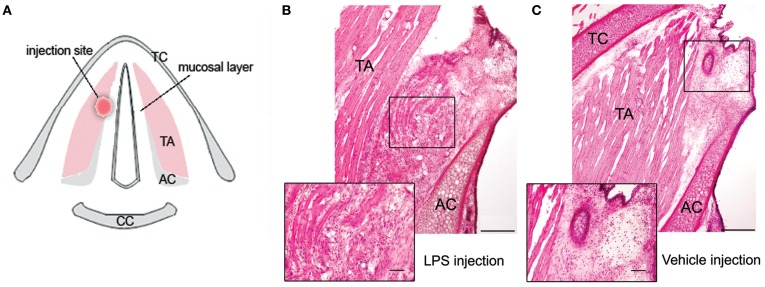
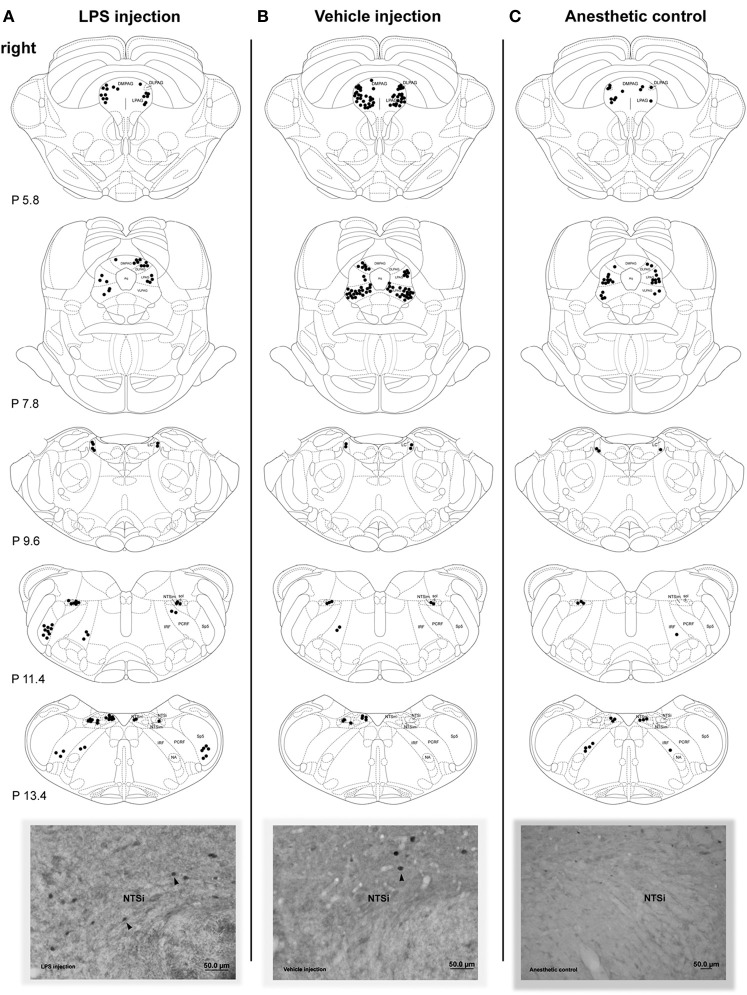
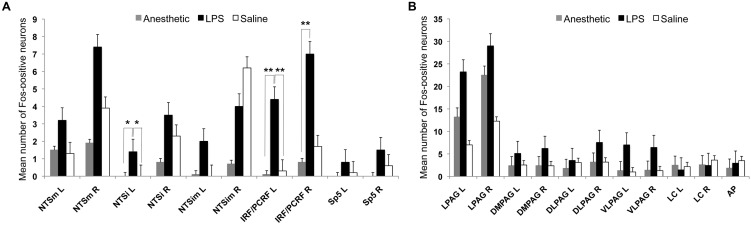
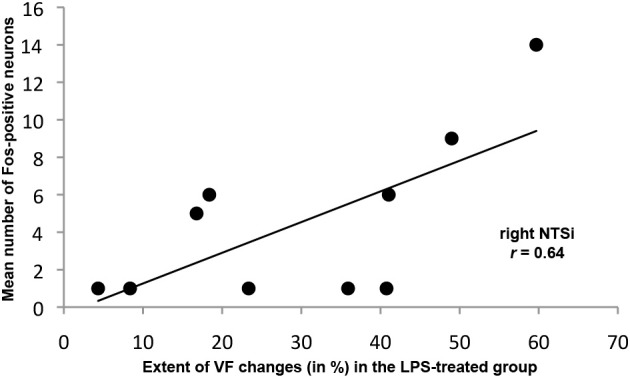
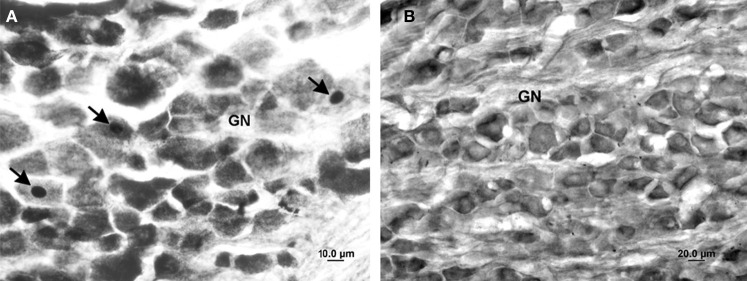
Somatosensory feedback from the larynx plays a critical role in regulation of normal upper airway functions, such as breathing, deglutition, and voice production, while altered laryngeal sensory feedback is known to elicit a variety of pathological reflex responses, including persistent coughing, dysphonia, and laryngospasm. Despite its clinical impact, the central mechanisms underlying the development of pathological laryngeal responses remain poorly understood. We examined the effects of persistent vocal fold (VF) inflammation and trauma, as frequent causes of long-lasting modulation of laryngeal sensory feedback, on brainstem immunoreactivity in the rat. Combined VF inflammation and trauma were induced by injection of lipopolysaccharide (LPS) solution and compared to VF trauma alone from injection of vehicle solution and to controls without any VF manipulations. Using a c-fos marker, we found significantly increased Fos-like immunoreactivity (FLI) in the bilateral intermediate/parvicellular reticular formation (IRF/PCRF) with a trend in the left solitary tract nucleus (NTS) only in animals with combined LPS-induced VF inflammation and trauma. Further, FLI in the right NTS was significantly correlated with the severity of LPS-induced VF changes. However, increased brainstem FLI response was not associated with FLI changes in the first-order neurons of the laryngeal afferents located in the nodose and jugular ganglia in either group. Our data indicate that complex VF alterations (i.e., inflammation/trauma vs. trauma alone) may cause prolonged excitability of the brainstem nuclei receiving a direct sensory input from the larynx, which, in turn, may lead to (mal)plastic changes within the laryngeal central sensory control.
Keywords: brainstem; immunoreactivity; inflammation; larynx; rat.
Figures
Similar articles
-
Descending Modulation of Laryngeal Vagal Sensory Processing in the Brainstem Orchestrated by the Submedius Thalamic Nucleus.J Neurosci. 2020 Dec 2;40(49):9426-9439. doi: 10.1523/JNEUROSCI.2430-20.2020. Epub 2020 Oct 28. J Neurosci. 2020. PMID: 33115928 Free PMC article.
-
Distribution of Fos-Like Immunoreactivity, Catecholaminergic and Serotoninergic Neurons Activated by the Laryngeal Chemoreflex in the Medulla Oblongata of Rats.PLoS One. 2015 Jun 18;10(6):e0130822. doi: 10.1371/journal.pone.0130822. eCollection 2015. PLoS One. 2015. PMID: 26087133 Free PMC article.
-
The Role of the Paratrigeminal Nucleus in Vagal Afferent Evoked Respiratory Reflexes: A Neuroanatomical and Functional Study in Guinea Pigs.Front Physiol. 2015 Dec 21;6:378. doi: 10.3389/fphys.2015.00378. eCollection 2015. Front Physiol. 2015. PMID: 26733874 Free PMC article.
-
Central integration of swallow and airway-protective reflexes.Am J Med. 2000 Mar 6;108 Suppl 4a:62S-67S. doi: 10.1016/s0002-9343(99)00340-x. Am J Med. 2000. PMID: 10718454 Review.
-
Brainstem viscerotopic organization of afferents and efferents involved in the control of swallowing.Am J Med. 2000 Mar 6;108 Suppl 4a:79S-86S. doi: 10.1016/s0002-9343(99)00343-5. Am J Med. 2000. PMID: 10718457 Review.
Cited by
-
Pathophysiology of Radiation-Induced Dysphagia in Head and Neck Cancer.Dysphagia. 2016 Jun;31(3):339-51. doi: 10.1007/s00455-016-9710-1. Epub 2016 Apr 20. Dysphagia. 2016. PMID: 27098922 Free PMC article. Review.
-
Activated Oxytocin Neurons in the PVN-DVC Pathway in Asthmatic Rats.Front Neuroanat. 2020 Jul 31;14:47. doi: 10.3389/fnana.2020.00047. eCollection 2020. Front Neuroanat. 2020. PMID: 32848637 Free PMC article.
-
The effect of combining action observation in virtual reality with kinesthetic motor imagery on cortical activity.Front Neurosci. 2023 Jun 13;17:1201865. doi: 10.3389/fnins.2023.1201865. eCollection 2023. Front Neurosci. 2023. PMID: 37383098 Free PMC article.
-
Sensory Innervation of the Larynx and the Search for Mucosal Mechanoreceptors.J Speech Lang Hear Res. 2021 Feb 17;64(2):371-391. doi: 10.1044/2020_JSLHR-20-00350. Epub 2021 Jan 19. J Speech Lang Hear Res. 2021. PMID: 33465318 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials