

The putative K(+) channel subunit AtKCO3 forms stable dimers in Arabidopsis
- PMID: 23162563
- PMCID: PMC3495302
- DOI: 10.3389/fpls.2012.00251
The putative K(+) channel subunit AtKCO3 forms stable dimers in Arabidopsis
Abstract
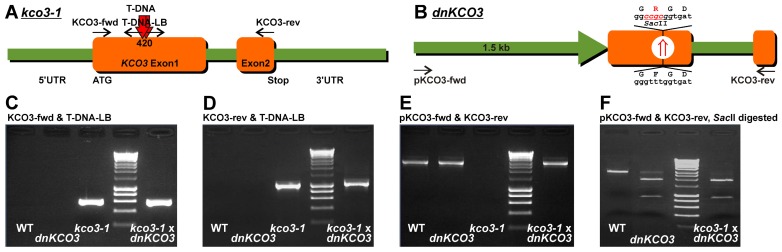
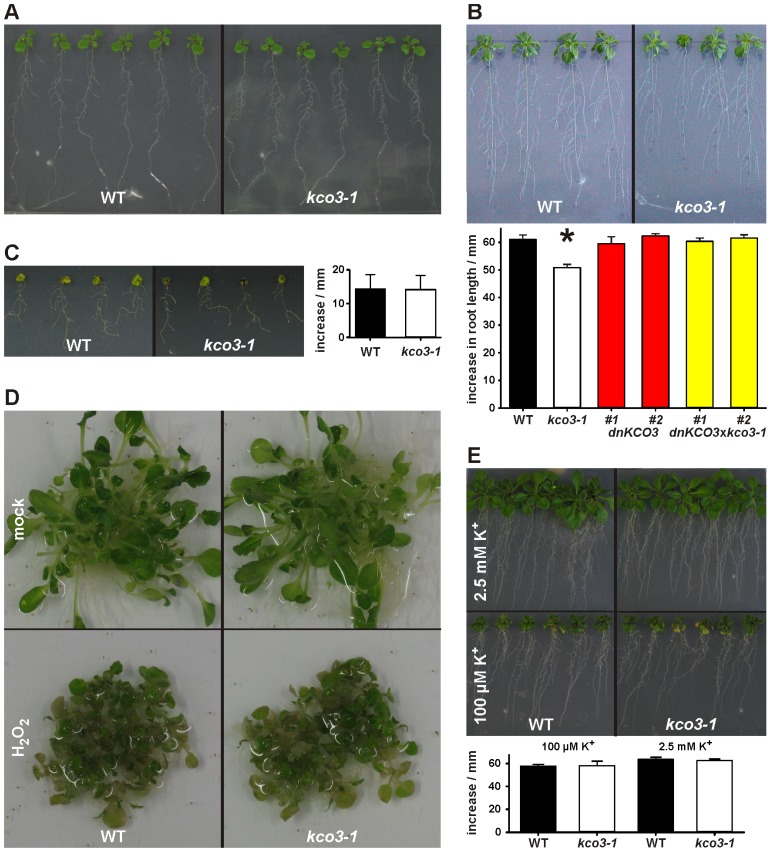
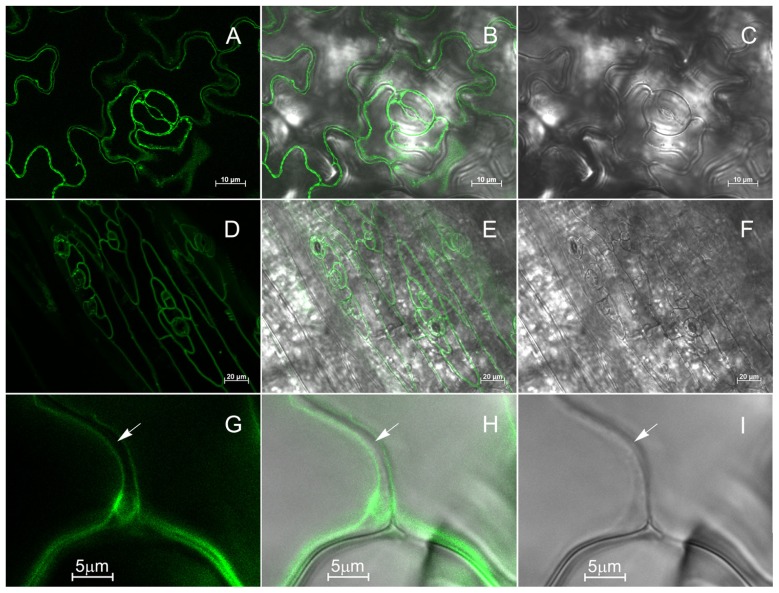
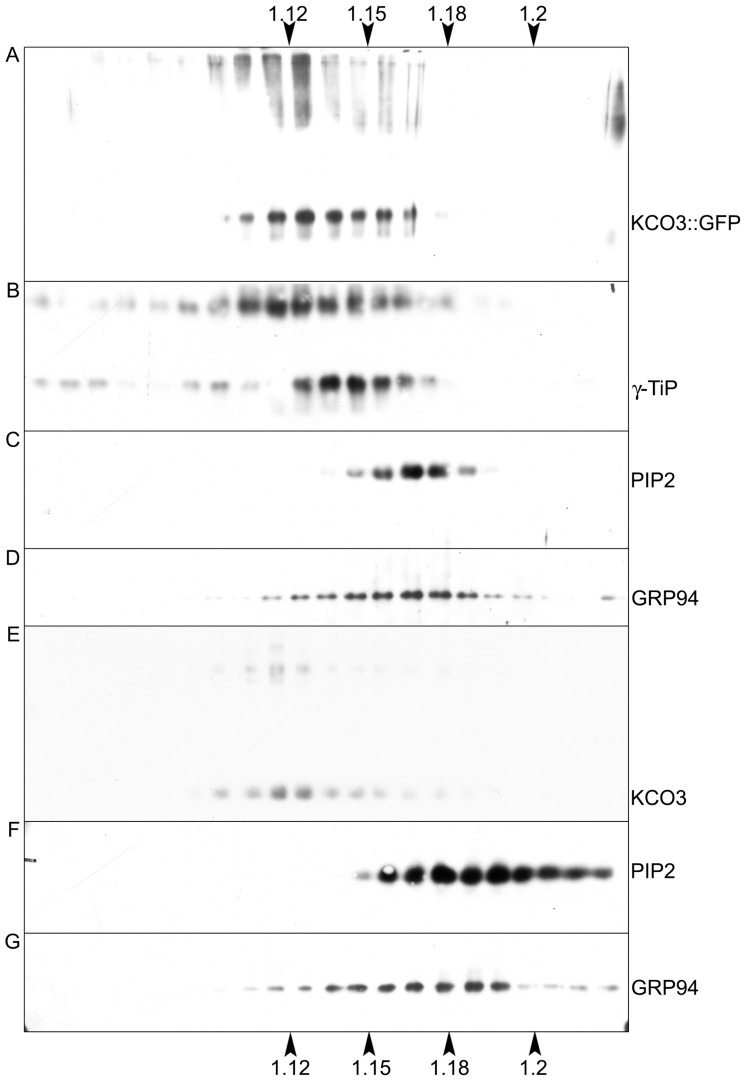
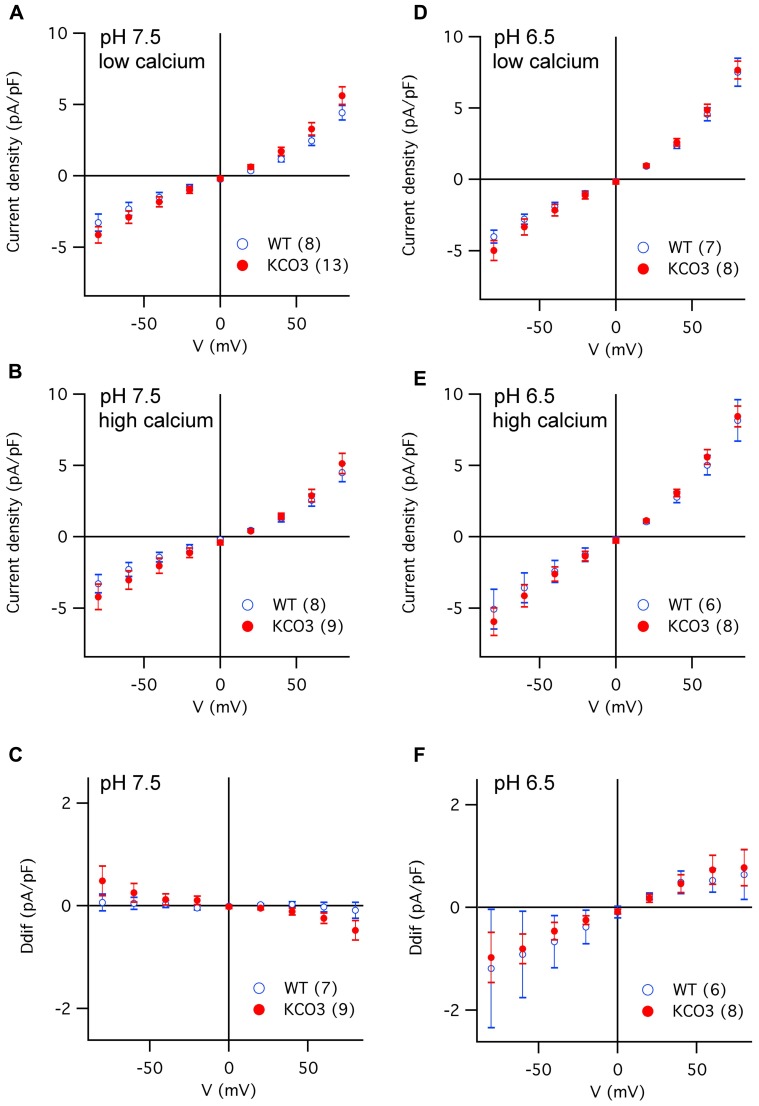
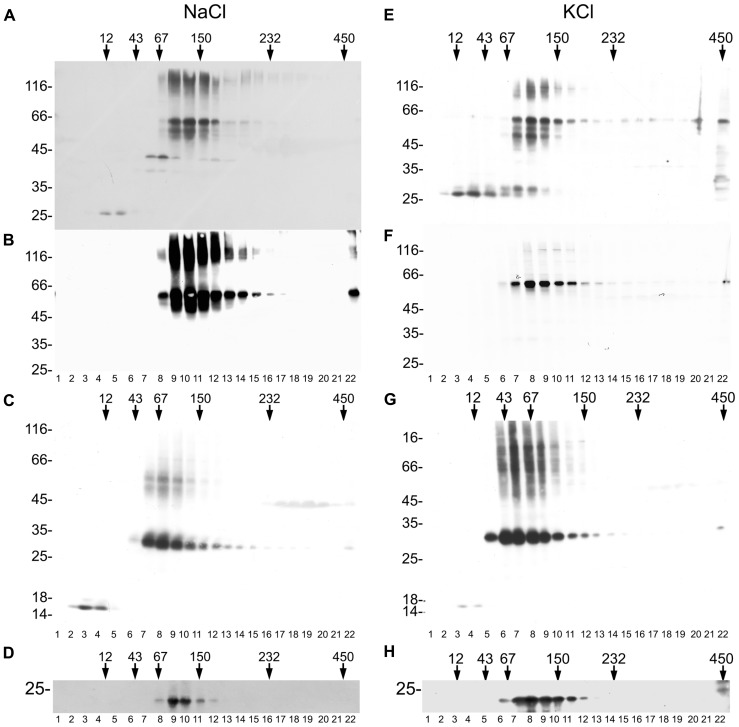
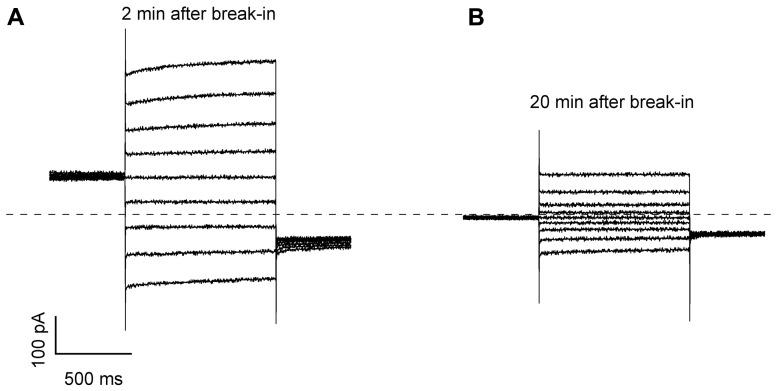
The permeation pore of K(+) channels is formed by four copies of the pore domain. AtKCO3 is the only putative voltage-independent K(+) channel subunit of Arabidopsis thaliana with a single pore domain. KCO3-like proteins recently emerged in evolution and, to date, have been found only in the genus Arabidopsis (A. thaliana and A. lyrata). We show that the absence of KCO3 does not cause marked changes in growth under various conditions. Only under osmotic stress we observed reduced root growth of the kco3-1 null-allele line. This phenotype was complemented by expressing a KCO3 mutant with an inactive pore, indicating that the function of KCO3 under osmotic stress does not depend on its direct ability to transport ions. Constitutively overexpressed AtKCO3 or AtKCO3::GFP are efficiently sorted to the tonoplast indicating that the protein is approved by the endoplasmic reticulum quality control. However, vacuoles isolated from transgenic plants do not have significant alterations in current density. Consistently, both AtKCO3 and AtKCO3::GFP are detected as homodimers upon velocity gradient centrifugation, an assembly state that would not allow for activity. We conclude that if AtKCO3 ever functions as a K(+) channel, active tetramers are held by particularly weak interactions, are formed only in unknown specific conditions and may require partner proteins.
Keywords: Arabidopsis; membrane proteins; potassium channels; protein assembly; tonoplast.
Figures
Similar articles
-
Analysis of Arabidopsis TPK2 and KCO3 reveals structural properties required for K+ channel function.Channels (Austin). 2020 Dec;14(1):336-346. doi: 10.1080/19336950.2020.1825894. Channels (Austin). 2020. PMID: 33016199 Free PMC article.
-
Members of the Arabidopsis AtTPK/KCO family form homomeric vacuolar channels in planta.Plant J. 2006 Oct;48(2):296-306. doi: 10.1111/j.1365-313X.2006.02868.x. Epub 2006 Sep 19. Plant J. 2006. PMID: 16984403
-
Roles of tandem-pore K+ channels in plants - a puzzle still to be solved.Plant Biol (Stuttg). 2010 Sep;12 Suppl 1:56-63. doi: 10.1111/j.1438-8677.2010.00353.x. Plant Biol (Stuttg). 2010. PMID: 20712621 Review.
-
Assembly and sorting of the tonoplast potassium channel AtTPK1 and its turnover by internalization into the vacuole.Plant Physiol. 2011 Aug;156(4):1783-96. doi: 10.1104/pp.111.177816. Epub 2011 Jun 22. Plant Physiol. 2011. PMID: 21697507 Free PMC article.
-
Recent updates on the physiology and evolution of plant TPK/KCO channels.Funct Plant Biol. 2023 Jan;50(1):17-28. doi: 10.1071/FP22117. Funct Plant Biol. 2023. PMID: 36220140 Review.
Cited by
-
In-Depth Genomic and Transcriptomic Analysis of Five K+ Transporter Gene Families in Soybean Confirm Their Differential Expression for Nodulation.Front Plant Sci. 2017 May 23;8:804. doi: 10.3389/fpls.2017.00804. eCollection 2017. Front Plant Sci. 2017. PMID: 28588592 Free PMC article.
-
The Membrane Transport System of the Guard Cell and Its Integration for Stomatal Dynamics.Plant Physiol. 2017 Jun;174(2):487-519. doi: 10.1104/pp.16.01949. Epub 2017 Apr 13. Plant Physiol. 2017. PMID: 28408539 Free PMC article. Review.
-
The Arabidopsis thaliana K+-Uptake Permease 5 (AtKUP5) Contains a Functional Cytosolic Adenylate Cyclase Essential for K+ Transport.Front Plant Sci. 2018 Nov 13;9:1645. doi: 10.3389/fpls.2018.01645. eCollection 2018. Front Plant Sci. 2018. PMID: 30483296 Free PMC article.
-
Identification of Two Auxin-Regulated Potassium Transporters Involved in Seed Maturation.Int J Mol Sci. 2018 Jul 22;19(7):2132. doi: 10.3390/ijms19072132. Int J Mol Sci. 2018. PMID: 30037141 Free PMC article.
-
Analysis of Arabidopsis TPK2 and KCO3 reveals structural properties required for K+ channel function.Channels (Austin). 2020 Dec;14(1):336-346. doi: 10.1080/19336950.2020.1825894. Channels (Austin). 2020. PMID: 33016199 Free PMC article.
References
-
- Allen G. J., Amtmann A., Sanders D. (1998). Calcium-dependent and calcium-independent K+ mobilization channels in Vicia faba guard cell vacuoles. J. Exp. Bot. 49 305–318
-
- Allen G. J., Sanders D. (1996). Control of ionic currents in guard cell vacuoles by cytosolic and luminal calcium. Plant J. 10 1055–1069 - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases