

Direct identification of rat iNKT cells reveals remarkable similarities to human iNKT cells and a profound deficiency in LEW rats
- PMID: 23165932
- PMCID: PMC4130812
- DOI: 10.1002/eji.201242565
Direct identification of rat iNKT cells reveals remarkable similarities to human iNKT cells and a profound deficiency in LEW rats
Abstract
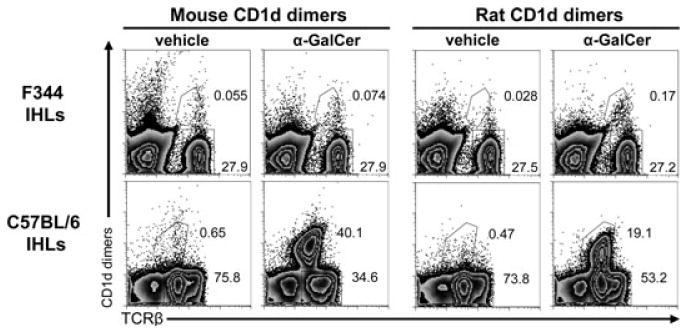
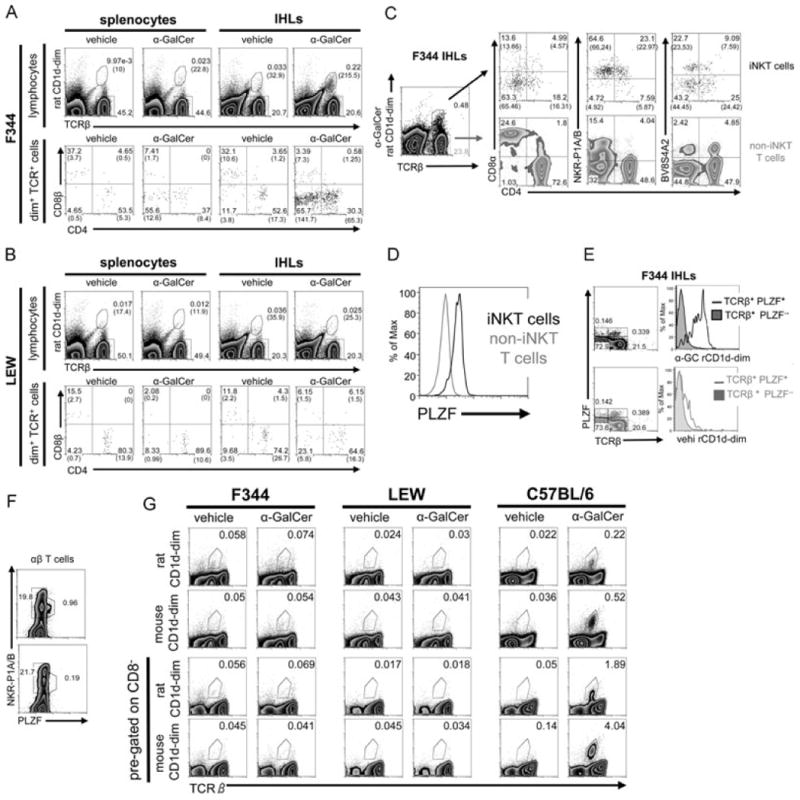
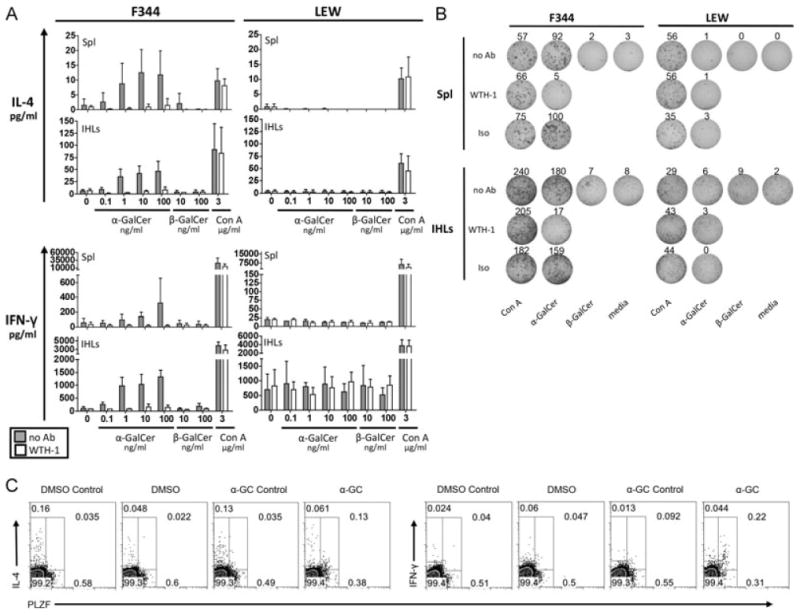
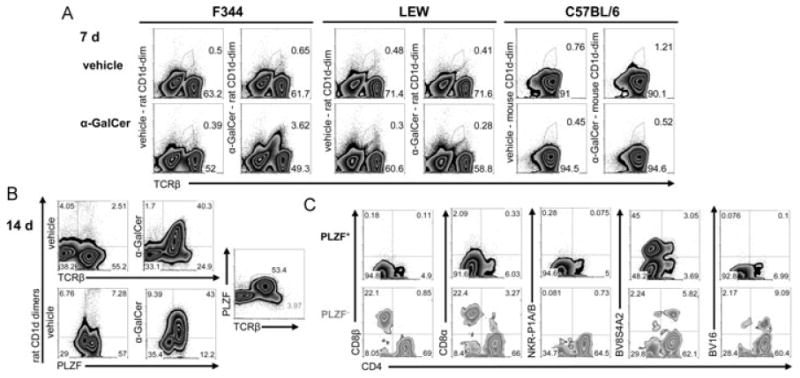
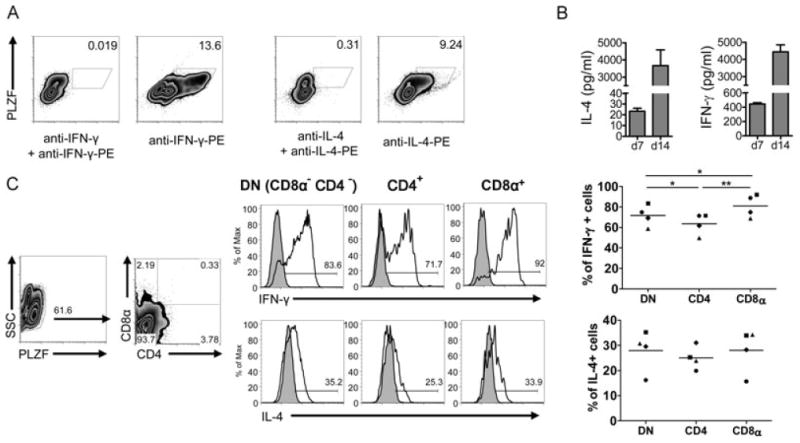
iNKT cells are a particular lymphocyte population with potent immunomodulatory capa-city; by promoting or suppressing immune responses against infections, tumors, and autoimmunity, iNKT cells are a promising target for immunotherapy. The hallmark of iNKT cells is the expression of a semiinvariant TCR (with an invariant α-chain comprising AV14 and AJ18 gene segments), which recognizes glycolipids presented by CD1d. Here, we identified iNKT cells for the first time in the rat using rat CD1d-dimers and PLZF staining. Importantly, in terms of frequencies (1.05% ± 0.52 SD of all intrahepatic αβ T cells), coreceptor expression and in vitro expansion features, iNKT cells from F344 inbred rats more closely resemble human iNKT cells than their mouse counterparts. In contrast, in LEW inbred rats, which are often used as models for organ-specific autoimmune diseases, iNKT cell numbers are near or below the detection limit. Interestingly, the usage of members of the rat AV14 gene family differed between F344 and LEW inbred rats. In conclusion, the similarities between F344 rat and human iNKT cells and the nearly absent iNKT cells in LEW rats make the rat a promising animal model for the study of iNKT cell-based therapies and of iNKT-cell biology.
© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
Similar articles
-
The hypervariable region 4 (HV4) and position 93 of the α chain modulate CD1d-glycolipid binding of iNKT TCRs.Eur J Immunol. 2015 Jul;45(7):2122-33. doi: 10.1002/eji.201545534. Epub 2015 May 12. Eur J Immunol. 2015. PMID: 25900449
-
The complementarity determining region 2 of BV8S2 (V beta 8.2) contributes to antigen recognition by rat invariant NKT cell TCR.J Immunol. 2006 Jun 15;176(12):7447-55. doi: 10.4049/jimmunol.176.12.7447. J Immunol. 2006. PMID: 16751390
-
Function and expression of CD1d and invariant natural killer T-cell receptor in the cotton rat (Sigmodon hispidus).Immunology. 2015 Dec;146(4):618-29. doi: 10.1111/imm.12532. Epub 2015 Oct 14. Immunology. 2015. PMID: 26346465 Free PMC article.
-
Invariant natural killer T cells: innate-like T cells with potent immunomodulatory activities.Tissue Antigens. 2009 Jun;73(6):535-45. doi: 10.1111/j.1399-0039.2009.01256.x. Epub 2009 Apr 8. Tissue Antigens. 2009. PMID: 19392798 Review.
-
Turned on by danger: activation of CD1d-restricted invariant natural killer T cells.Immunology. 2012 Sep;137(1):20-7. doi: 10.1111/j.1365-2567.2012.03612.x. Immunology. 2012. PMID: 22734667 Free PMC article. Review.
Cited by
-
Species Specific Differences of CD1d Oligomer Loading In Vitro.PLoS One. 2015 Nov 24;10(11):e0143449. doi: 10.1371/journal.pone.0143449. eCollection 2015. PLoS One. 2015. PMID: 26599805 Free PMC article.
-
Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition).Eur J Immunol. 2019 Oct;49(10):1457-1973. doi: 10.1002/eji.201970107. Eur J Immunol. 2019. PMID: 31633216 Free PMC article.
-
Guidelines for the use of flow cytometry and cell sorting in immunological studies (third edition).Eur J Immunol. 2021 Dec;51(12):2708-3145. doi: 10.1002/eji.202170126. Epub 2021 Dec 7. Eur J Immunol. 2021. PMID: 34910301 Free PMC article. Review.
-
Polymorphisms in the CD1d promoter that regulate CD1d gene expression are associated with impaired NKT cell development.J Immunol. 2014 Jan 1;192(1):189-99. doi: 10.4049/jimmunol.1301451. Epub 2013 Dec 4. J Immunol. 2014. PMID: 24307737 Free PMC article.
-
The Vγ9Vδ2 T Cell Antigen Receptor and Butyrophilin-3 A1: Models of Interaction, the Possibility of Co-Evolution, and the Case of Dendritic Epidermal T Cells.Front Immunol. 2014 Dec 19;5:648. doi: 10.3389/fimmu.2014.00648. eCollection 2014. Front Immunol. 2014. PMID: 25566259 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
