

The role of UNC93B1 protein in surface localization of TLR3 receptor and in cell priming to nucleic acid agonists
- PMID: 23166319
- PMCID: PMC3537042
- DOI: 10.1074/jbc.M112.413922
The role of UNC93B1 protein in surface localization of TLR3 receptor and in cell priming to nucleic acid agonists
Abstract
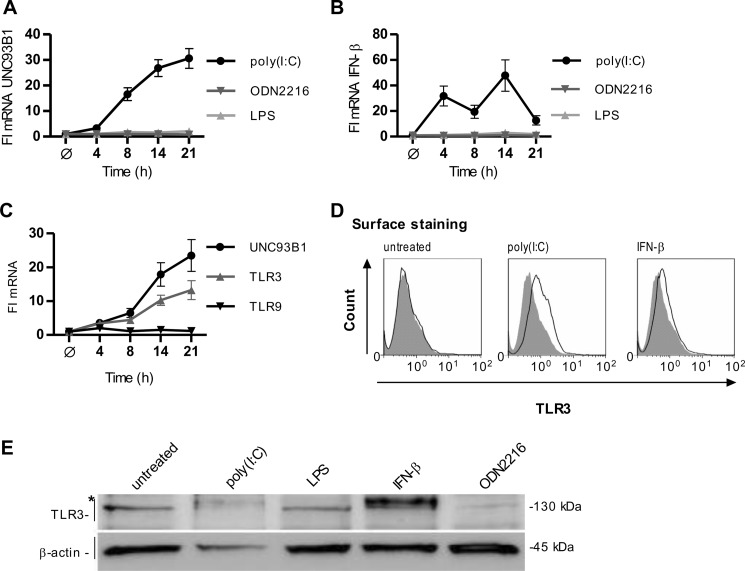
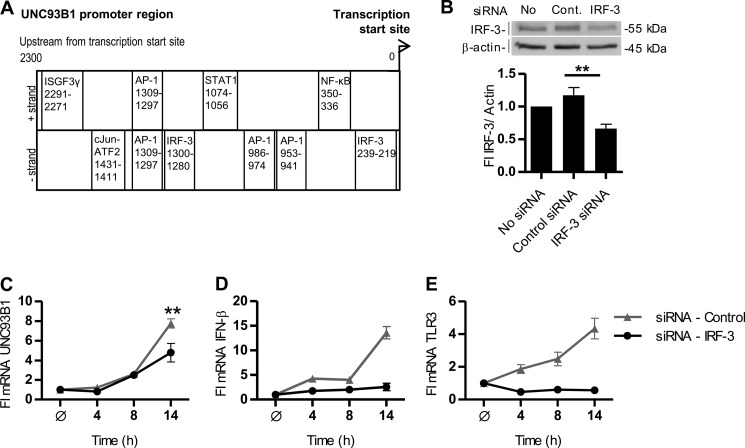
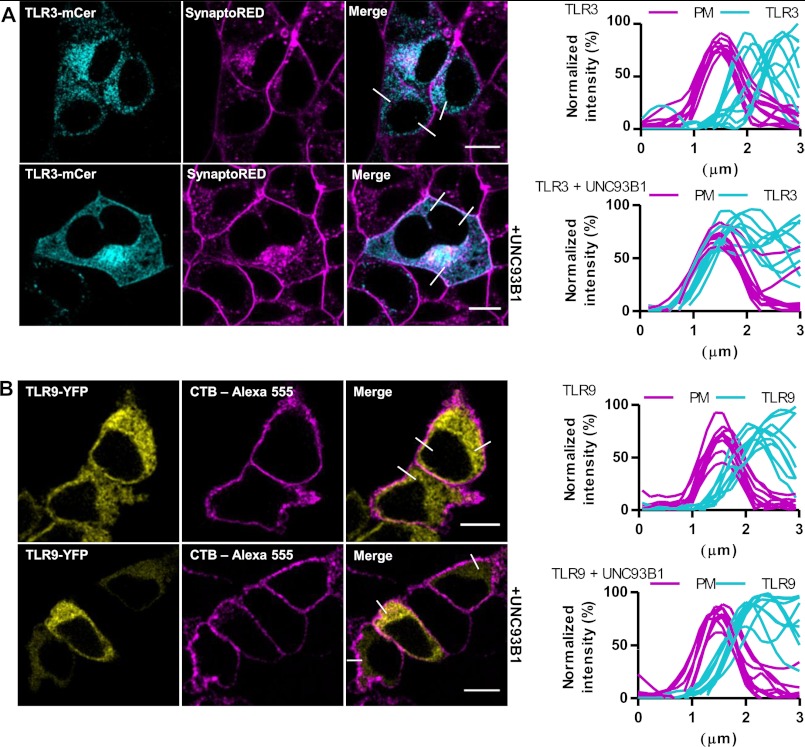
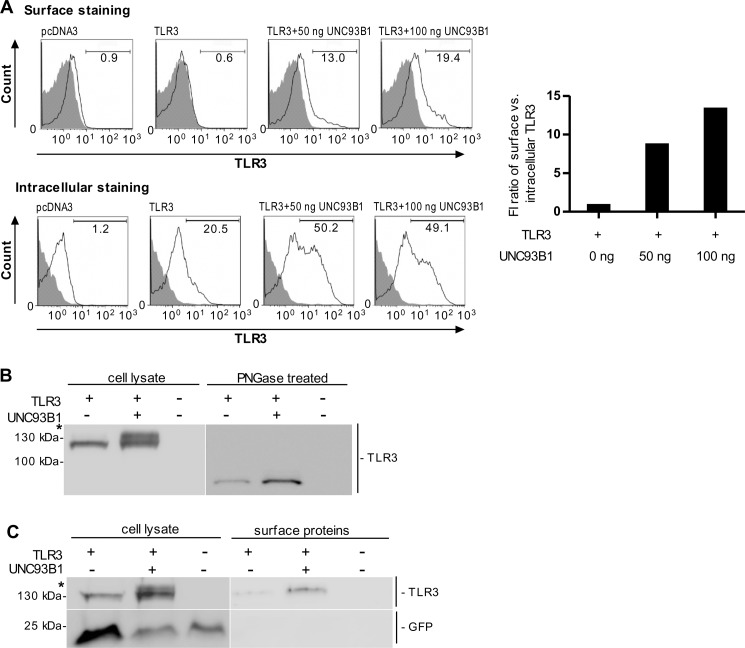
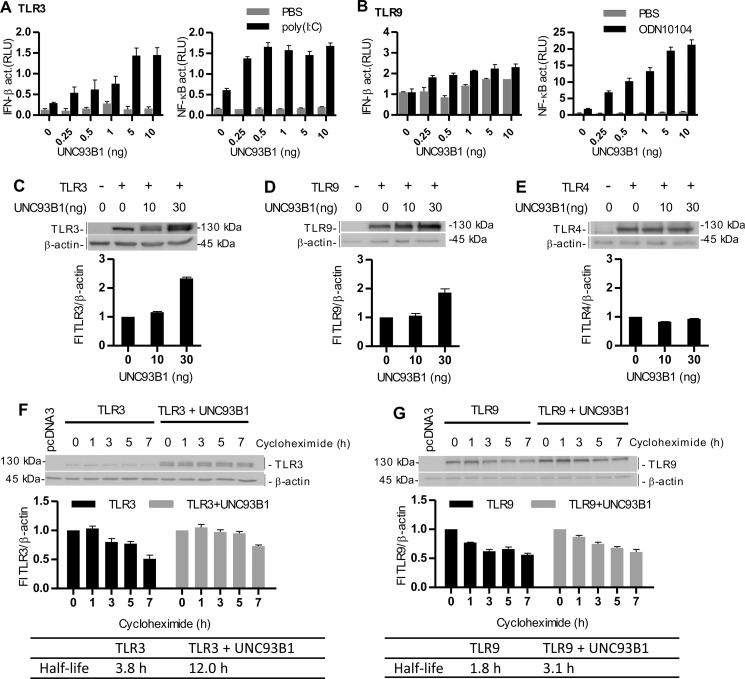
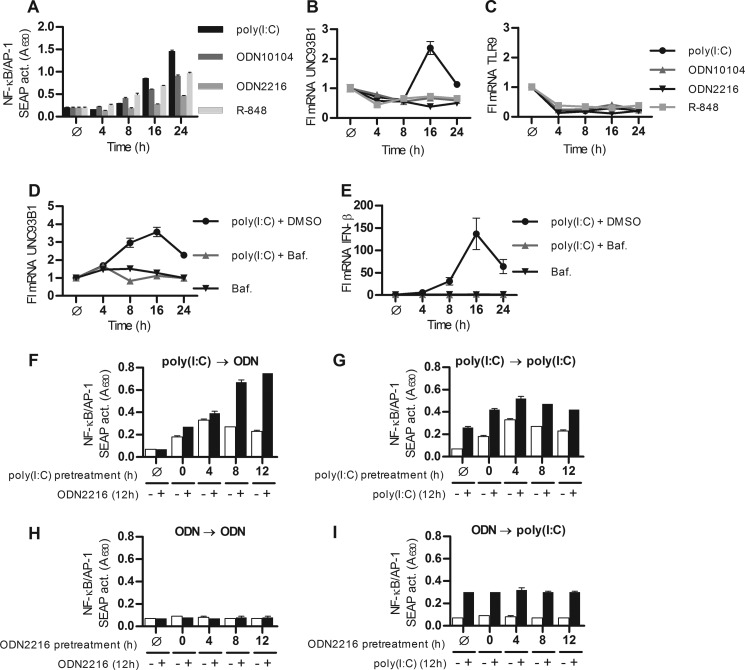
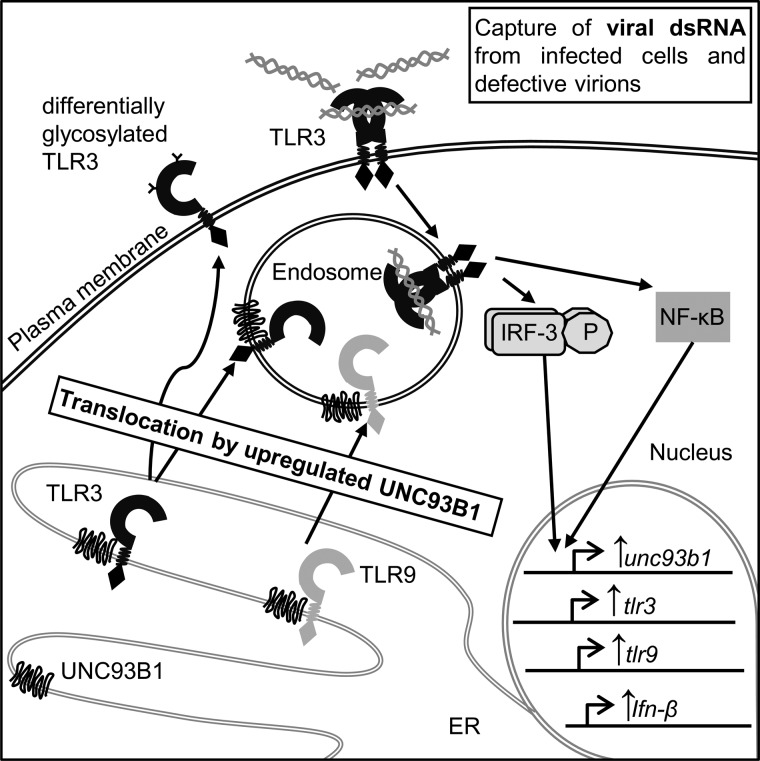
Translocation of nucleic acid-sensing (NAS) Toll-like receptors (TLRs) to endosomes is essential for response to microbial nucleic acids as well as for prevention of the autoimmune response. The accessory protein UNC93B1 is indispensable for activation of NAS TLRs because it regulates their response through trafficking to endosomes. We observed that poly(I:C) up-regulates transcription of UNC93B1 and promotes trafficking of TLR3 to the plasma membrane in human epithelial cell line. Up-regulation of UNC93B1 is triggered through TLR3 activation by poly(I:C). Further studies revealed that expression of UNC93B1 promotes trafficking of differentially glycosylated TLR3, but not other NAS TLRs, to the plasma membrane. UNC93B1 promoter region contains binding sites for poly(I:C)- and type I interferon-inducible regulatory elements. UNC93B1 also increases the protein lifetime of TLR3 and TLR9 and augments signaling of all NAS TLRs. Furthermore, we discovered that poly(I:C) pretreatment primes B-cells to the activation by ssDNA via up-regulation of UNC93B1. Our findings identified TLR3 as the important regulator of UNC93B1 that in turn governs the responsiveness of all NAS TLRs.
Figures
Similar articles
-
The ectodomain of TLR3 receptor is required for its plasma membrane translocation.PLoS One. 2014 Mar 20;9(3):e92391. doi: 10.1371/journal.pone.0092391. eCollection 2014. PLoS One. 2014. PMID: 24651829 Free PMC article.
-
Acidic amino acid residues in the juxtamembrane region of the nucleotide-sensing TLRs are important for UNC93B1 binding and signaling.J Immunol. 2013 May 15;190(10):5287-95. doi: 10.4049/jimmunol.1202767. Epub 2013 Apr 12. J Immunol. 2013. PMID: 23585677
-
UNC93B1 and nucleic acid-sensing Toll-like receptors mediate host resistance to infection with Leishmania major.J Biol Chem. 2013 Mar 8;288(10):7127-36. doi: 10.1074/jbc.M112.407684. Epub 2013 Jan 16. J Biol Chem. 2013. PMID: 23325805 Free PMC article.
-
Cell Surface Expression of Endosomal Toll-Like Receptors-A Necessity or a Superfluous Duplication?Front Immunol. 2021 Feb 1;11:620972. doi: 10.3389/fimmu.2020.620972. eCollection 2020. Front Immunol. 2021. PMID: 33597952 Free PMC article. Review.
-
Dynamic control of nucleic-acid-sensing Toll-like receptors by the endosomal compartment.Int Immunol. 2021 Nov 25;33(12):835-840. doi: 10.1093/intimm/dxab037. Int Immunol. 2021. PMID: 34223897 Review.
Cited by
-
Combined TLR-3/TLR-8 Signaling in the Presence of α-Type-1 Cytokines Represents a Novel and Potent Dendritic Cell Type-1, Anti-Cancer Maturation Protocol.Cells. 2022 Feb 28;11(5):835. doi: 10.3390/cells11050835. Cells. 2022. PMID: 35269457 Free PMC article.
-
Immunity and Coagulation in COVID-19.Int J Mol Sci. 2024 Oct 19;25(20):11267. doi: 10.3390/ijms252011267. Int J Mol Sci. 2024. PMID: 39457048 Free PMC article. Review.
-
Expression Profiles of Long Noncoding RNA and mRNA in Epicardial Adipose Tissue in Patients with Heart Failure.Biomed Res Int. 2019 Jul 4;2019:3945475. doi: 10.1155/2019/3945475. eCollection 2019. Biomed Res Int. 2019. PMID: 31355260 Free PMC article.
-
New application of anti-TLR monoclonal antibodies: detection, inhibition and protection.Inflamm Regen. 2018 Jul 3;38:11. doi: 10.1186/s41232-018-0068-7. eCollection 2018. Inflamm Regen. 2018. PMID: 29988708 Free PMC article. Review.
-
Nucleic Acid Immunity.Adv Immunol. 2017;133:121-169. doi: 10.1016/bs.ai.2016.11.001. Epub 2016 Dec 15. Adv Immunol. 2017. PMID: 28215278 Free PMC article.
References
-
- Akira S., Takeda K. (2004) Toll-like receptor signalling. Nat. Rev. Immunol. 4, 499–511 - PubMed
-
- Beutler B. (2004) Innate immunity. An overview. Mol. Immunol. 40, 845–859 - PubMed
-
- McGettrick A. F., O'Neill L. A. (2010) Localisation and trafficking of Toll-like receptors. An important mode of regulation. Curr. Opin. Immunol. 22, 20–27 - PubMed
-
- Choe J., Kelker M. S., Wilson I. A. (2005) Crystal structure of human toll-like receptor 3 (TLR3) ectodomain. Science 309, 581–585 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
