

Molecular interaction studies of HIV-1 matrix protein p17 and heparin: identification of the heparin-binding motif of p17 as a target for the development of multitarget antagonists
- PMID: 23166320
- PMCID: PMC3542999
- DOI: 10.1074/jbc.M112.400077
Molecular interaction studies of HIV-1 matrix protein p17 and heparin: identification of the heparin-binding motif of p17 as a target for the development of multitarget antagonists
Abstract
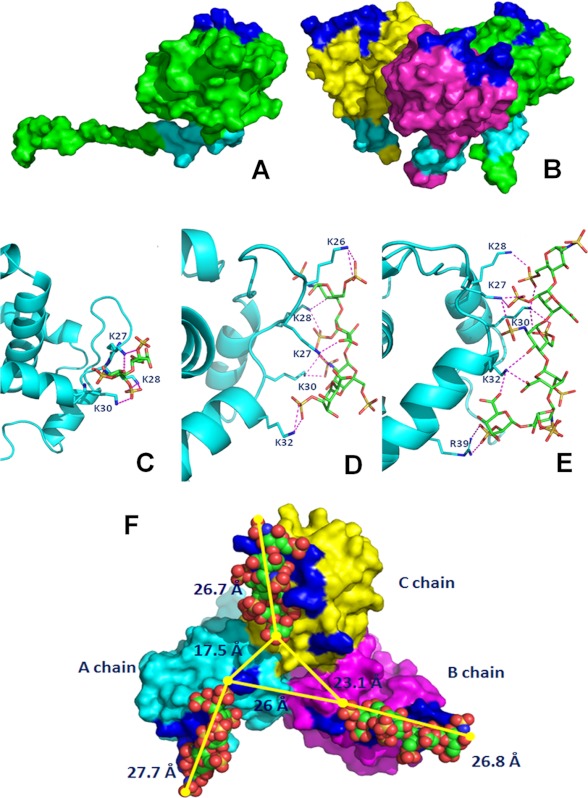
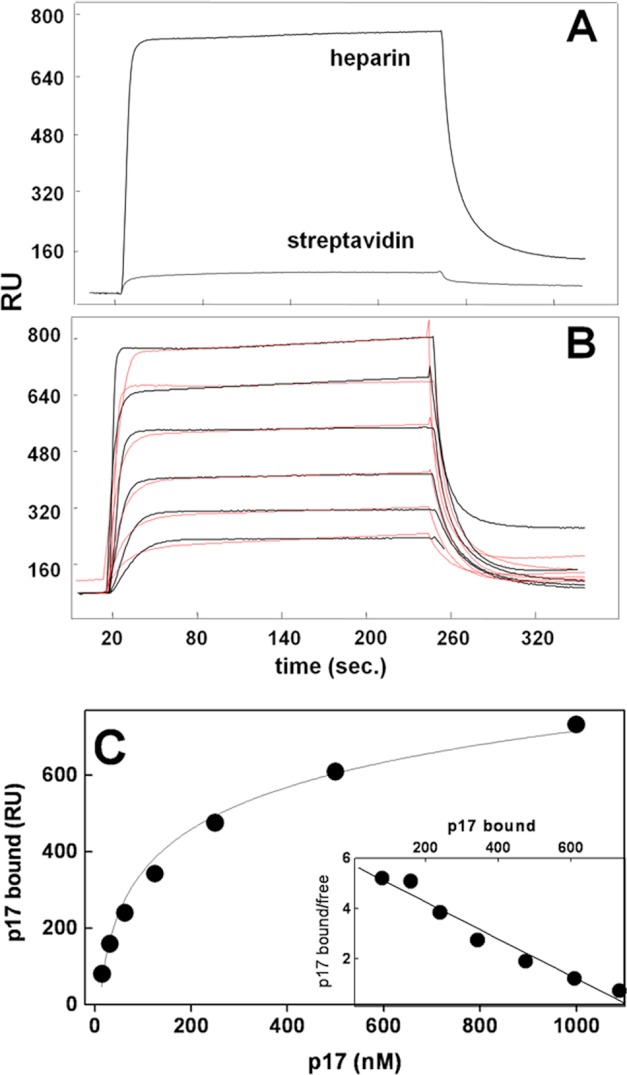
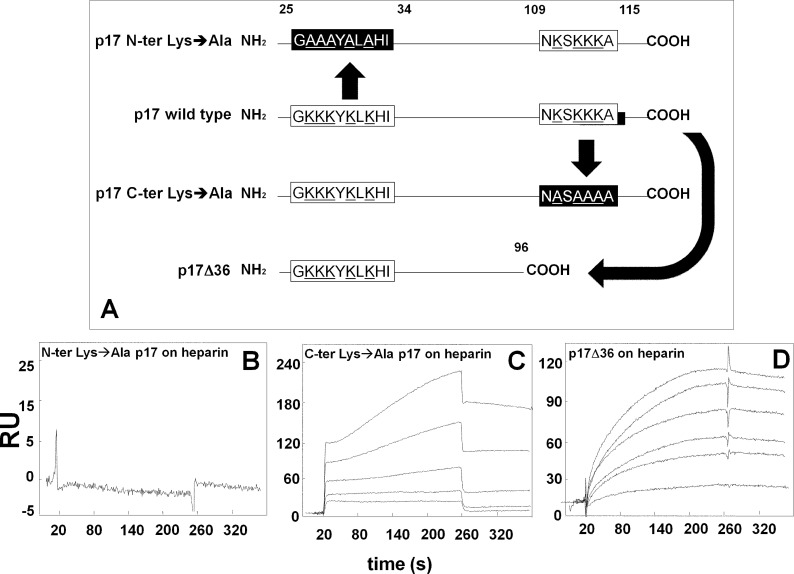
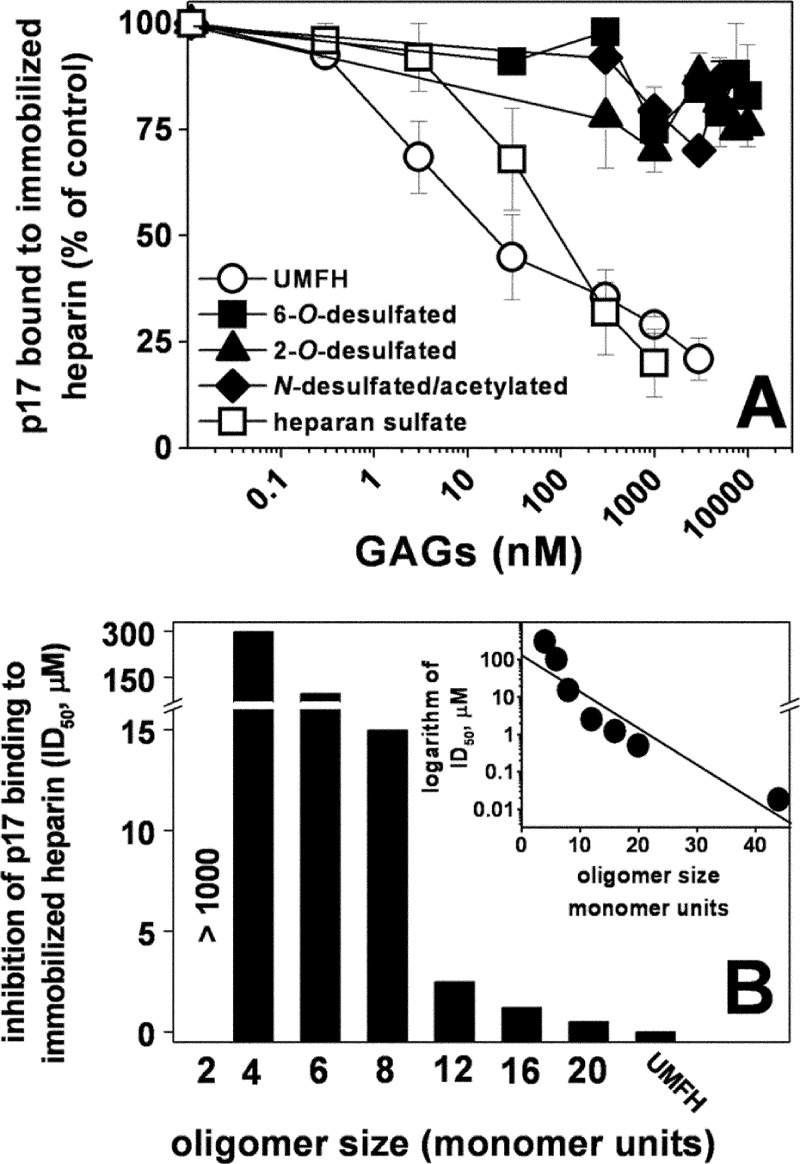
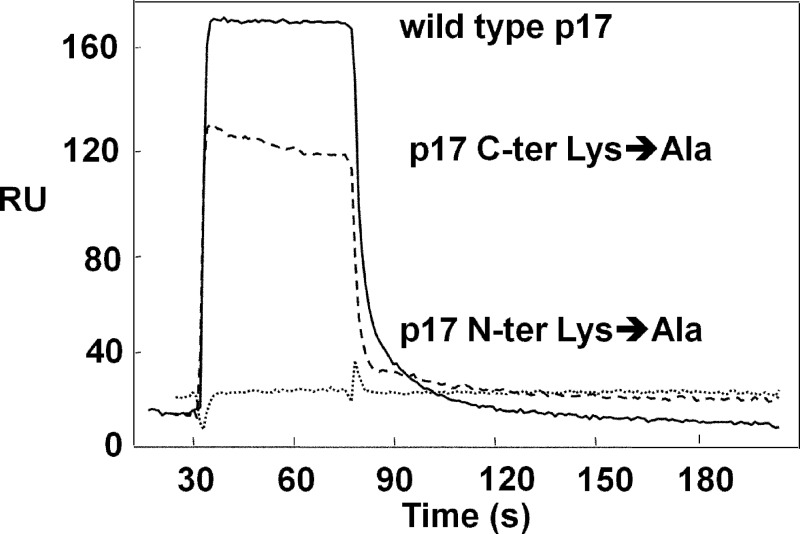
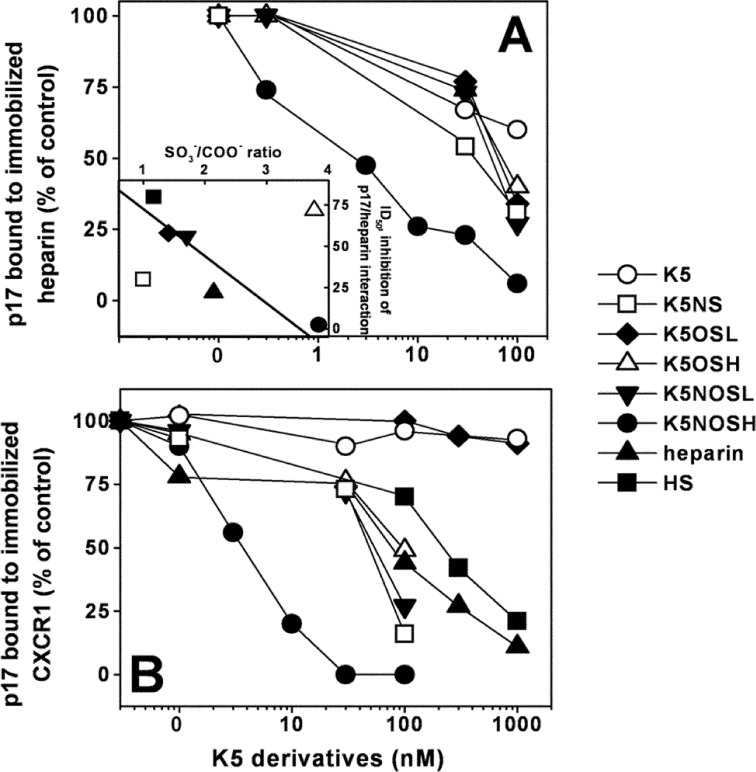
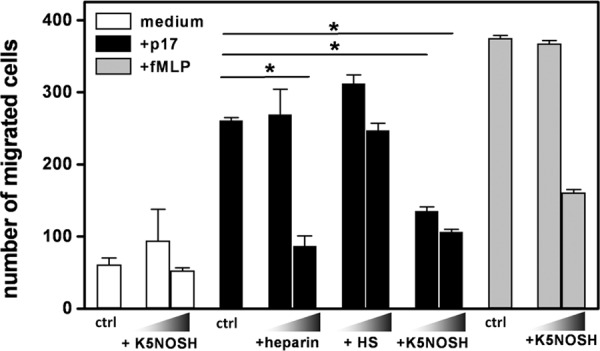
Once released by HIV(+) cells, p17 binds heparan sulfate proteoglycans (HSPGs) and CXCR1 on leukocytes causing their dysfunction. By exploiting an approach integrating computational modeling, site-directed mutagenesis of p17, chemical desulfation of heparin, and surface plasmon resonance, we characterized the interaction of p17 with heparin, a HSPG structural analog, and CXCR1. p17 binds to heparin with an affinity (K(d) = 190 nm) that is similar to those of other heparin-binding viral proteins. Two stretches of basic amino acids (basic motifs) are present in p17 N and C termini. Neutralization (Arg→Ala substitution) of the N-terminal, but not of the C-terminal basic motif, causes the loss of p17 heparin-binding capacity. The N-terminal heparin-binding motif of p17 partially overlaps the CXCR1-binding domain. Accordingly, its neutralization prevents also p17 binding to the chemochine receptor. Competition experiments demonstrated that free heparin and heparan sulfate (HS), but not selectively 2-O-, 6-O-, and N-O desulfated heparins, prevent p17 binding to substrate-immobilized heparin, indicating that the sulfate groups of the glycosaminoglycan mediate p17 interaction. Evaluation of the p17 antagonist activity of a panel of biotechnological heparins derived by chemical sulfation of the Escherichia coli K5 polysaccharide revealed that the highly N,O-sulfated derivative prevents the binding of p17 to both heparin and CXCR1, thus inhibiting p17-driven chemotactic migration of human monocytes with an efficiency that is higher than those of heparin and HS. Here, we characterized at a molecular level the interaction of p17 with its cellular receptors, laying the basis for the development of heparin-mimicking p17 antagonists.
Figures
References
-
- Fiorentini S., Riboldi E., Facchetti F., Avolio M., Fabbri M., Tosti G., Becker P. D., Guzman C. A., Sozzani S., Caruso A. (2008) HIV-1 matrix protein p17 induces human plasmacytoid dendritic cells to acquire a migratory immature cell phenotype. Proc. Natl. Acad. Sci. U.S.A. 105, 3867–3872 - PMC - PubMed
-
- Budka H. (1990) Human immunodeficiency virus (HIV) envelope and core proteins in CNS tissues of patients with the acquired immune deficiency syndrome (AIDS). Acta Neuropathol. 79, 611–619 - PubMed
-
- Popovic M., Tenner-Racz K., Pelser C., Stellbrink H. J., van Lunzen J., Lewis G., Kalyanaraman V. S., Gallo R. C., Racz P. (2005) Persistence of HIV-1 structural proteins and glycoproteins in lymph nodes of patients under highly active antiretroviral therapy. Proc. Natl. Acad. Sci. U.S.A. 102, 14807–14812 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
