

Functional expression of SCL/TAL1 interrupting locus (Stil) protects retinal dopaminergic cells from neurotoxin-induced degeneration
- PMID: 23166330
- PMCID: PMC3543037
- DOI: 10.1074/jbc.M112.417089
Functional expression of SCL/TAL1 interrupting locus (Stil) protects retinal dopaminergic cells from neurotoxin-induced degeneration
Abstract
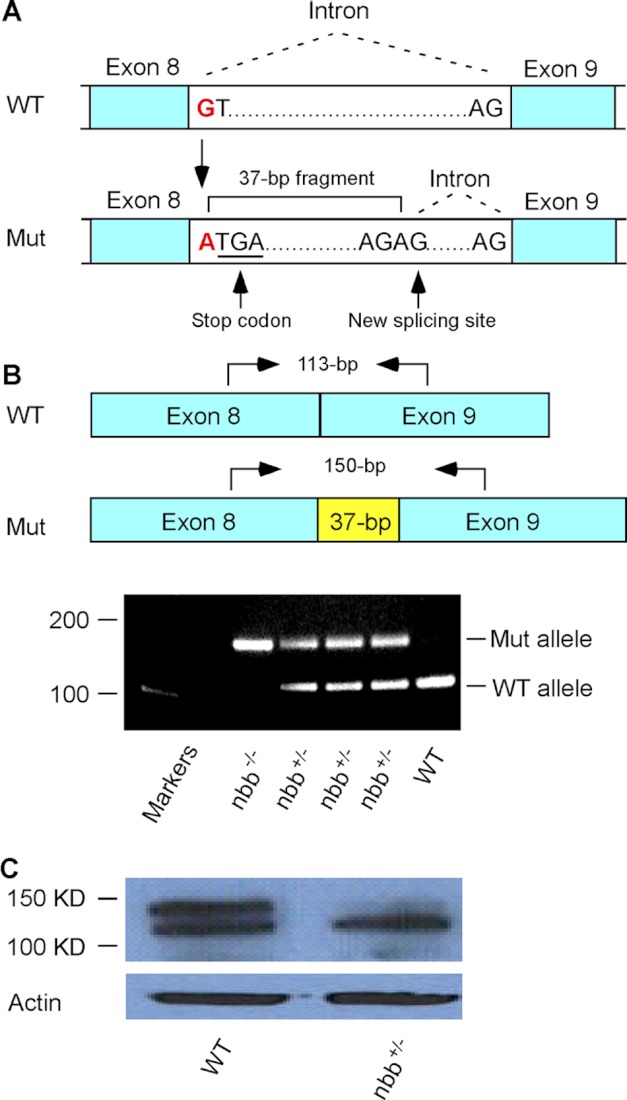
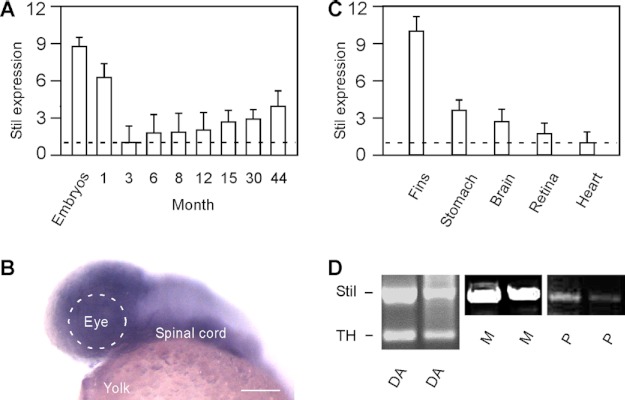
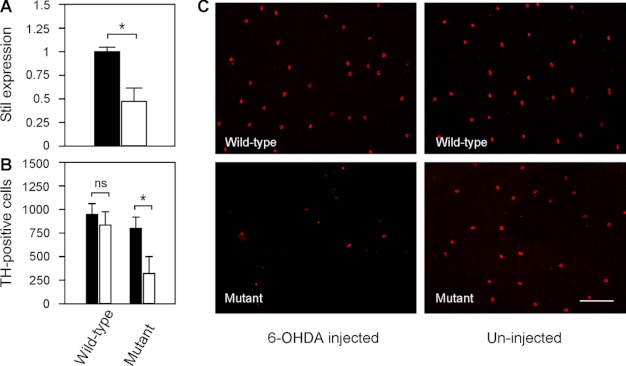
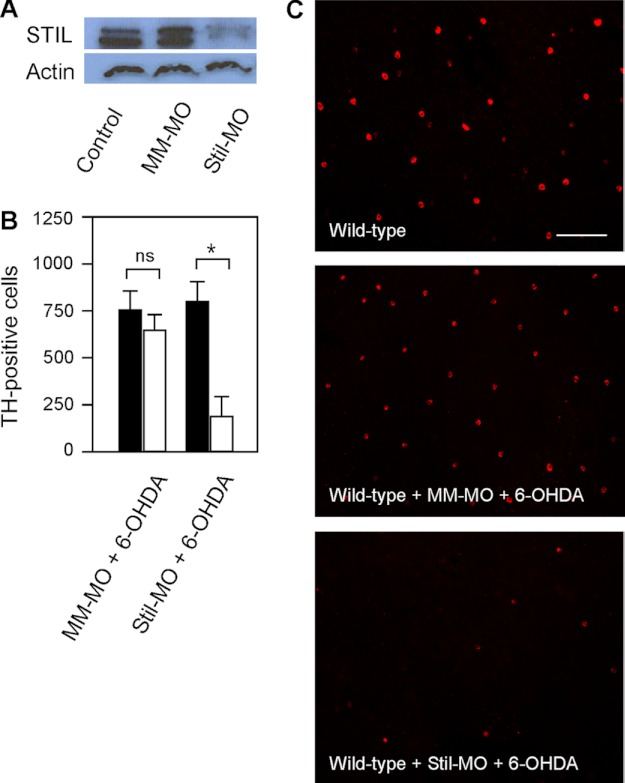
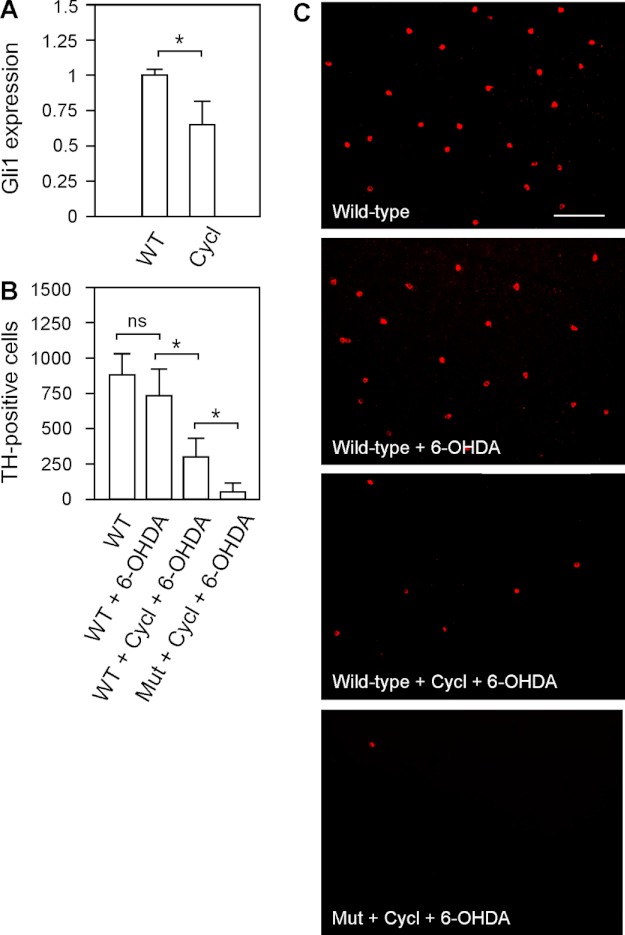
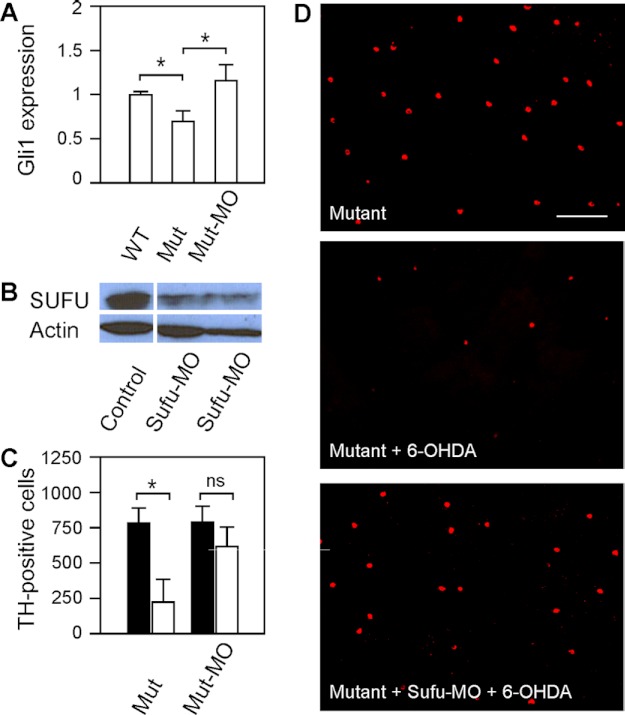
We previously isolated a dominant mutation, night blindness b (nbb), which causes a late onset of retinal dopaminergic cell degeneration in zebrafish. In this study, we cloned the zebrafish nbb locus. Sequencing results revealed that nbb is a homolog of the vertebrate SCL/TAL1 interrupting locus (Stil). The Stil gene has been shown to play important roles in the regulation of vertebrate embryonic neural development and human cancer cell proliferation. In this study, we demonstrate that functional expression of Stil is also required for neural survival. In zebrafish, decreased expression of Stil resulted in increased toxic susceptibility of retinal dopaminergic cells to 6-hydroxydopamine. Increases in Stil-mediated Shh signaling transduction (i.e. by knocking down the Shh repressor Sufu) prevented dopaminergic cell death induced by neurotoxic insult. The data suggest that the oncogene Stil also plays important roles in neural protection.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
