

Mitotic chromosomes are compacted laterally by KIF4 and condensin and axially by topoisomerase IIα
- PMID: 23166350
- PMCID: PMC3514791
- DOI: 10.1083/jcb.201202155
Mitotic chromosomes are compacted laterally by KIF4 and condensin and axially by topoisomerase IIα
Abstract
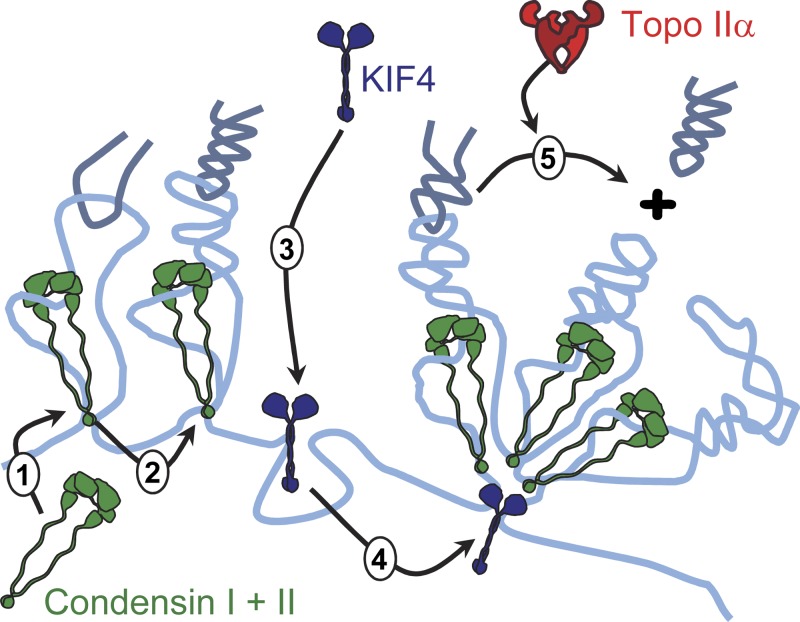
Mitotic chromosome formation involves a relatively minor condensation of the chromatin volume coupled with a dramatic reorganization into the characteristic "X" shape. Here we report results of a detailed morphological analysis, which revealed that chromokinesin KIF4 cooperated in a parallel pathway with condensin complexes to promote the lateral compaction of chromatid arms. In this analysis, KIF4 and condensin were mutually dependent for their dynamic localization on the chromatid axes. Depletion of either caused sister chromatids to expand and compromised the "intrinsic structure" of the chromosomes (defined in an in vitro assay), with loss of condensin showing stronger effects. Simultaneous depletion of KIF4 and condensin caused complete loss of chromosome morphology. In these experiments, topoisomerase IIα contributed to shaping mitotic chromosomes by promoting the shortening of the chromatid axes and apparently acting in opposition to the actions of KIF4 and condensins. These three proteins are major determinants in shaping the characteristic mitotic chromosome morphology.
Figures









Comment in
-
Chromosomes: getting the architecture right.Nat Rev Mol Cell Biol. 2013 Jan;14(1):2-3. doi: 10.1038/nrm3498. Epub 2012 Dec 12. Nat Rev Mol Cell Biol. 2013. PMID: 23232562 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
