

The quorum sensing volatile molecule 2-amino acetophenon modulates host immune responses in a manner that promotes life with unwanted guests
- PMID: 23166496
- PMCID: PMC3499575
- DOI: 10.1371/journal.ppat.1003024
The quorum sensing volatile molecule 2-amino acetophenon modulates host immune responses in a manner that promotes life with unwanted guests
Abstract
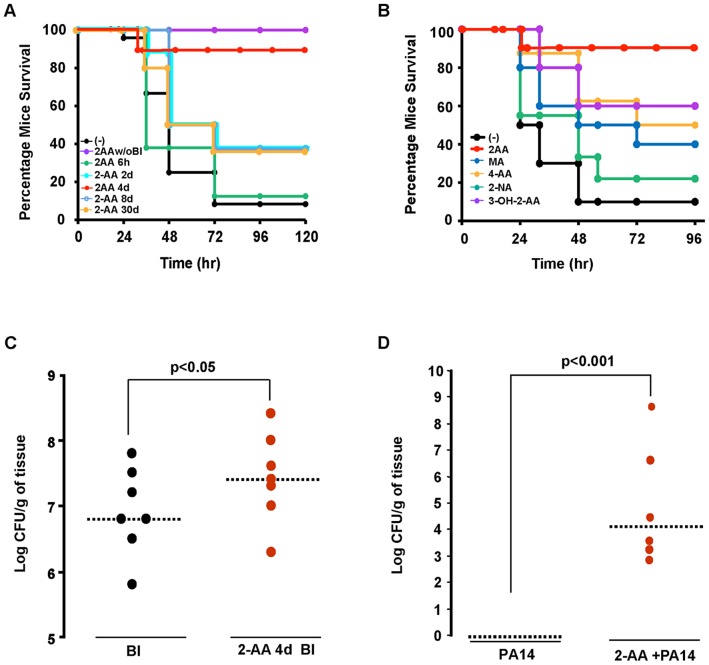
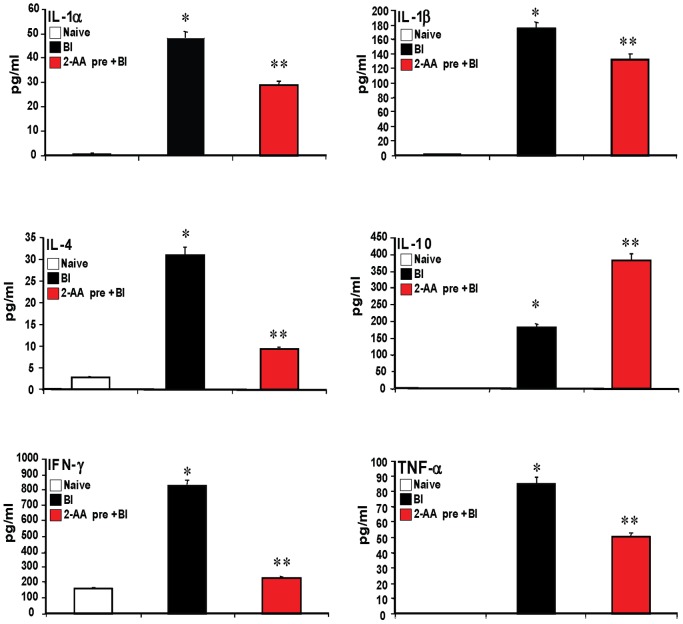
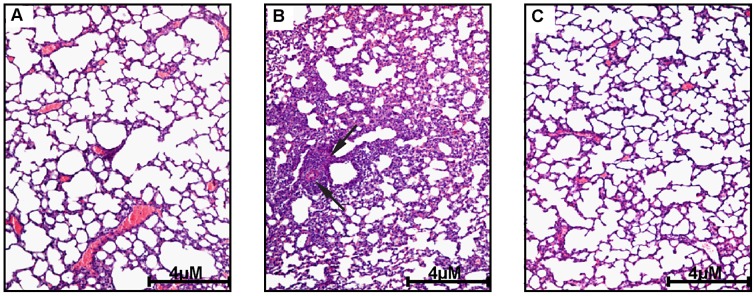
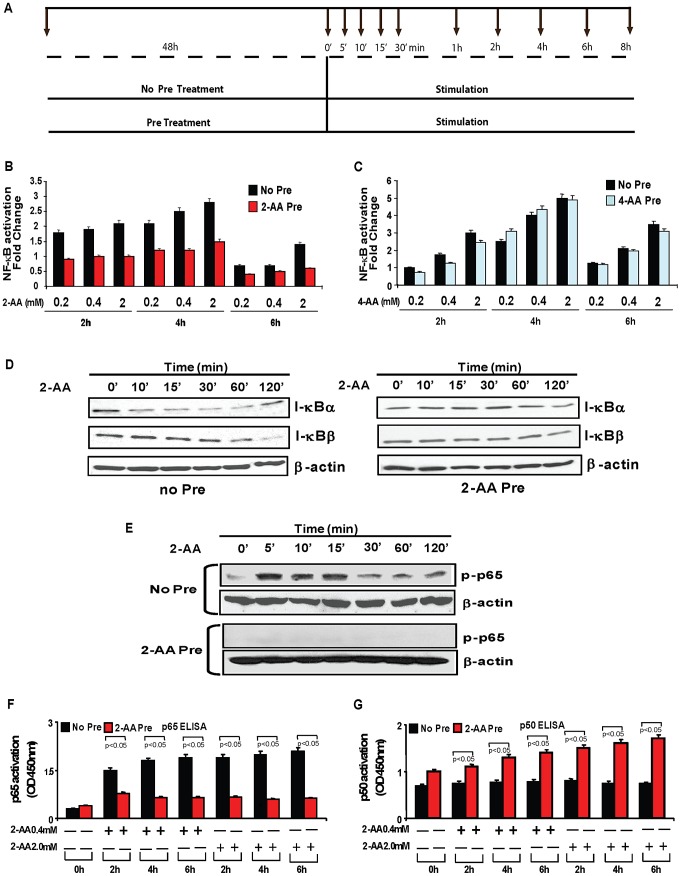
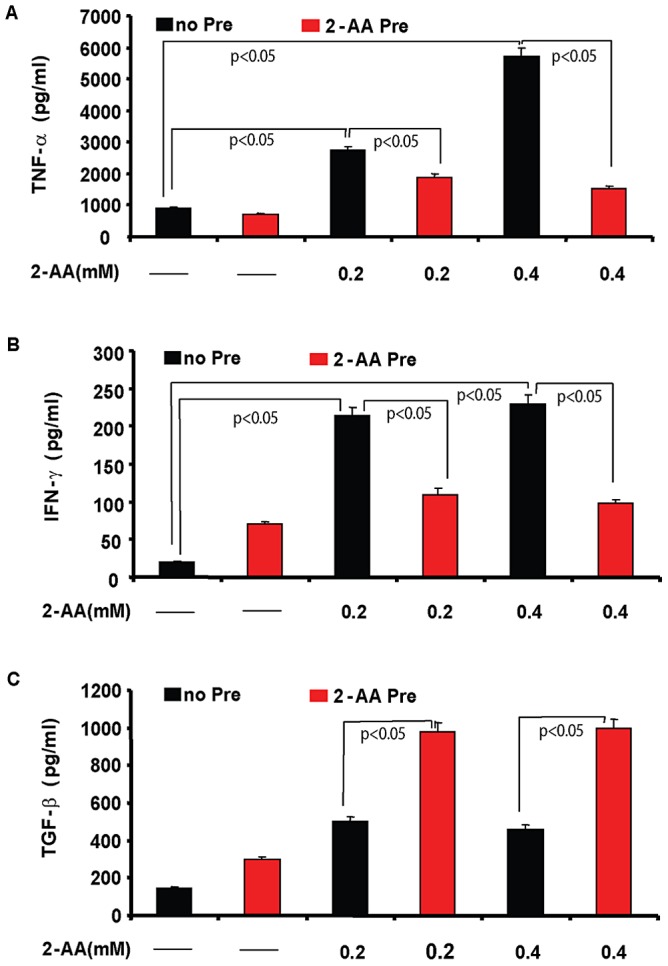
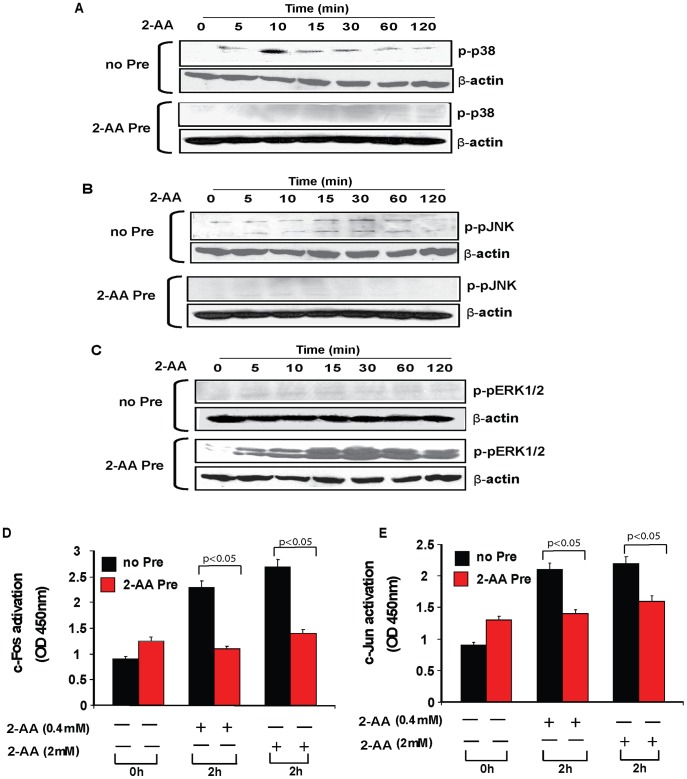
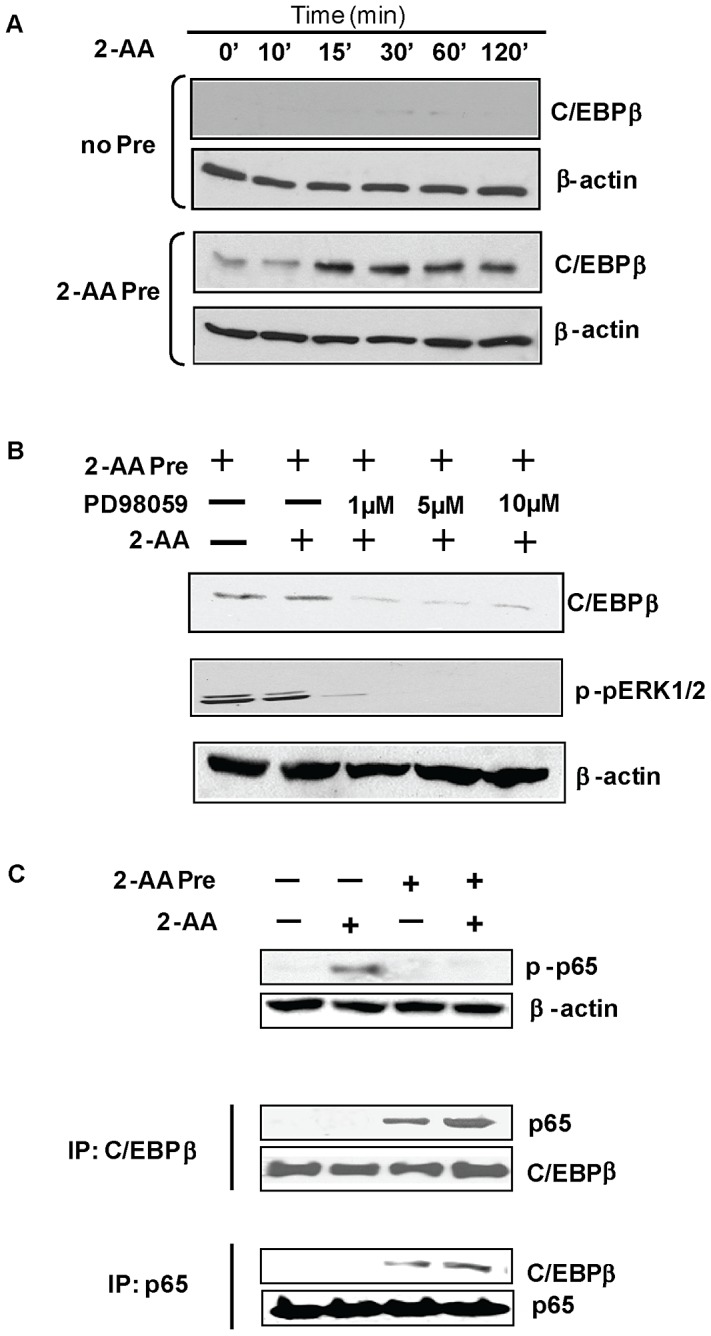
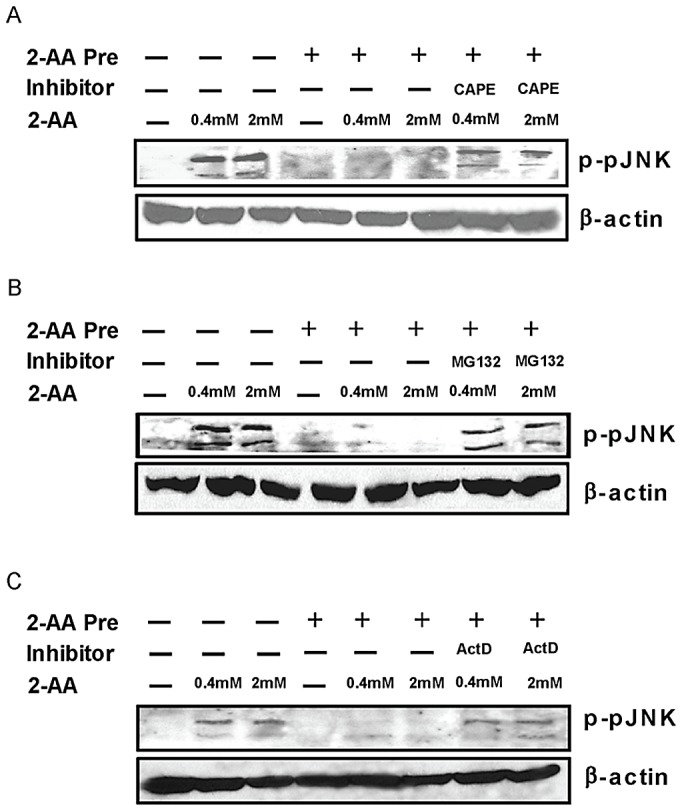
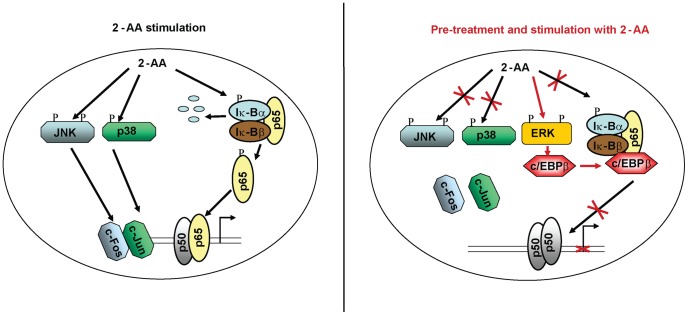
Increasing evidence indicates that bacterial quorum sensing (QS) signals are important mediators of immunomodulation. However, whether microbes utilize these immunomodulatory signals to maintain infection remain unclear. Here, we show that the Pseudomonas aeruginosa QS-regulated molecule 2-amino acetophenone (2-AA) modulates host immune responses in a manner that increases host ability to cope with this pathogen. Mice treated with 2-AA prior to infection had a 90% survival compared to 10% survival rate observed in the non-pretreated infected mice. Whilst 2-AA stimulation activates key innate immune response pathways involving mitogen-activated protein kinases (MAPKs), nuclear factor (NF)-κB, and pro-inflammatory cytokines, it attenuates immune response activation upon pretreatment, most likely by upregulating anti-inflammatory cytokines. 2-AA host pretreatment is characterized by a transcriptionally regulated block of c-JUN N-terminal kinase (JNK) and NF-κB activation, with relatively preserved activation of extracellular regulated kinase (ERK) 1/2. These kinase changes lead to CCAAT/enhancer-binding protein-β (c/EBPβ) activation and formation of the c/EBPβ-p65 complex that prevents NF-κB activation. 2-AA's aptitude for dampening the inflammatory processes while increasing host survival and pathogen persistence concurs with its ability to signal bacteria to switch to a chronic infection mode. Our results reveal a QS immunomodulatory signal that promotes original aspects of interkingdom communication. We propose that this communication facilitates pathogen persistence, while enabling host tolerance to infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Antunes LC, Ferreira RB, Buckner MM, Finlay BB (2010) Quorum sensing in bacterial virulence. Microbiology 156: 2271–2282. - PubMed
-
- Keller L, Surette MG (2006) Communication in bacteria: an ecological and evolutionary perspective. Nat Rev Microbiol 4: 249–258. - PubMed
-
- Deziel E, Gopalan S, Tampakaki AP, Lepine F, Padfield KE, et al. (2005) The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-L-homoserine lactones. Mol Microbiol 55: 998–1014. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
