

Sensitivity of mitochondrial transcription and resistance of RNA polymerase II dependent nuclear transcription to antiviral ribonucleosides
- PMID: 23166498
- PMCID: PMC3499576
- DOI: 10.1371/journal.ppat.1003030
Sensitivity of mitochondrial transcription and resistance of RNA polymerase II dependent nuclear transcription to antiviral ribonucleosides
Abstract
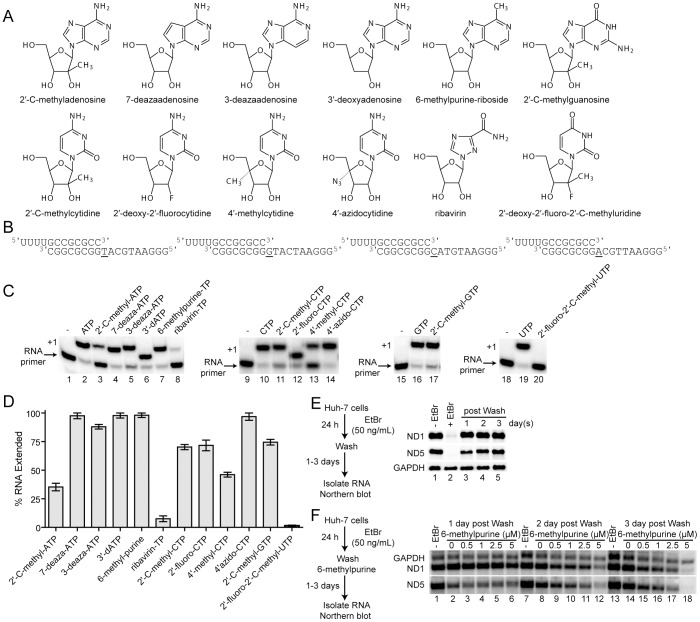
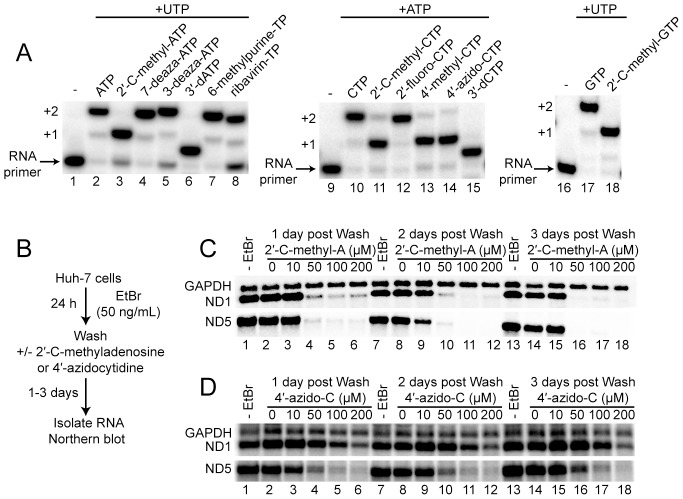
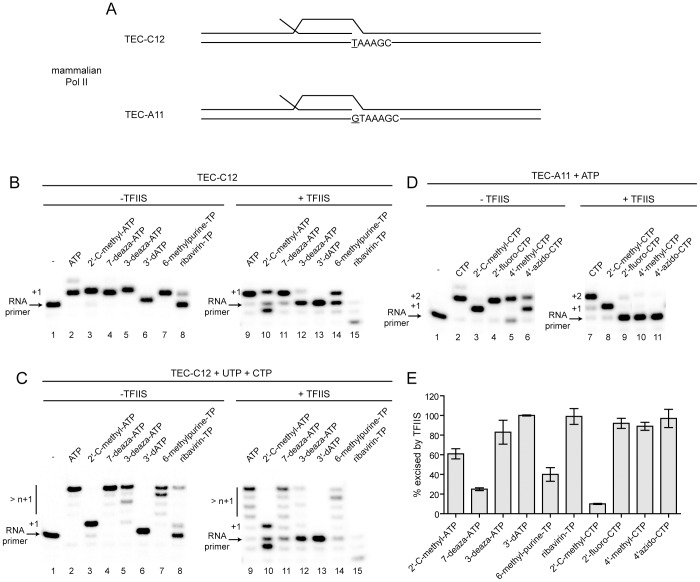
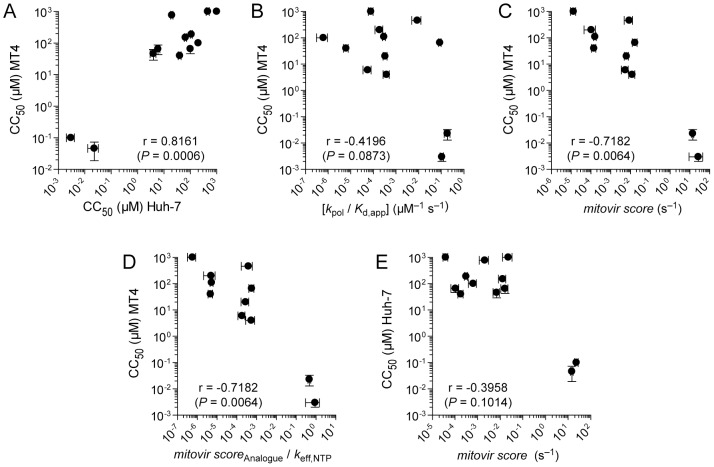
Ribonucleoside analogues have potential utility as anti-viral, -parasitic, -bacterial and -cancer agents. However, their clinical applications have been limited by off target effects. Development of antiviral ribonucleosides for treatment of hepatitis C virus (HCV) infection has been hampered by appearance of toxicity during clinical trials that evaded detection during preclinical studies. It is well established that the human mitochondrial DNA polymerase is an off target for deoxyribonucleoside reverse transcriptase inhibitors. Here we test the hypothesis that triphosphorylated metabolites of therapeutic ribonucleoside analogues are substrates for cellular RNA polymerases. We have used ribonucleoside analogues with activity against HCV as model compounds for therapeutic ribonucleosides. We have included ribonucleoside analogues containing 2'-C-methyl, 4'-methyl and 4'-azido substituents that are non-obligate chain terminators of the HCV RNA polymerase. We show that all of the anti-HCV ribonucleoside analogues are substrates for human mitochondrial RNA polymerase (POLRMT) and eukaryotic core RNA polymerase II (Pol II) in vitro. Unexpectedly, analogues containing 2'-C-methyl, 4'-methyl and 4'-azido substituents were inhibitors of POLRMT and Pol II. Importantly, the proofreading activity of TFIIS was capable of excising these analogues from Pol II transcripts. Evaluation of transcription in cells confirmed sensitivity of POLRMT to antiviral ribonucleosides, while Pol II remained predominantly refractory. We introduce a parameter termed the mitovir (mitochondrial dysfunction caused by antiviral ribonucleoside) score that can be readily obtained during preclinical studies that quantifies the mitochondrial toxicity potential of compounds. We suggest the possibility that patients exhibiting adverse effects during clinical trials may be more susceptible to damage by nucleoside analogs because of defects in mitochondrial or nuclear transcription. The paradigm reported here should facilitate development of ribonucleosides with a lower potential for toxicity.
Conflict of interest statement
JYF, ASR, AC, JP, JEV, YP, YX, YT, DB, OB and WZ are current or former employees and shareholders of Gilead Sciences, Inc. that is responsible for the active clinical development of GS-7977 and has a partnership with Roche related to the development of mericitabine. JJA and CEC are collaborating with INDIGO Biosciences, Inc. for the development and distribution of mitochondrial toxicity assays. This does not alter our adherence to all PLOS Pathogens policies on sharing data and materials.
Figures
References
-
- Braaten D, editor(1975) Chemistry, Biology, and Clinical Use of Nucleoside Analogs. 3–610 p.
-
- Carrasco L, Vazquez D (1984) Molecular bases for the action and selectivity of nucleoside antibiotics. Med Res Rev 4: 471–512. - PubMed
-
- Fox JJ, Watanabe KA, Bloch A (1966) Nucleoside antibiotics. Prog Nucleic Acid Res Mol Biol 5: 251–313. - PubMed
-
- Brown NA (2009) Progress towards improving antiviral therapy for hepatitis C with hepatitis C virus polymerase inhibitors. Part I: Nucleoside analogues. Expert Opin Investig Drugs 18: 709–725. - PubMed
-
- Lech-Maranda E, Korycka A, Robak T (2006) Pharmacological and clinical studies on purine nucleoside analogs–new anticancer agents. Mini Rev Med Chem 6: 575–581. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
