

Disruption of mouse Cenpj, a regulator of centriole biogenesis, phenocopies Seckel syndrome
- PMID: 23166506
- PMCID: PMC3499256
- DOI: 10.1371/journal.pgen.1003022
Disruption of mouse Cenpj, a regulator of centriole biogenesis, phenocopies Seckel syndrome
Abstract
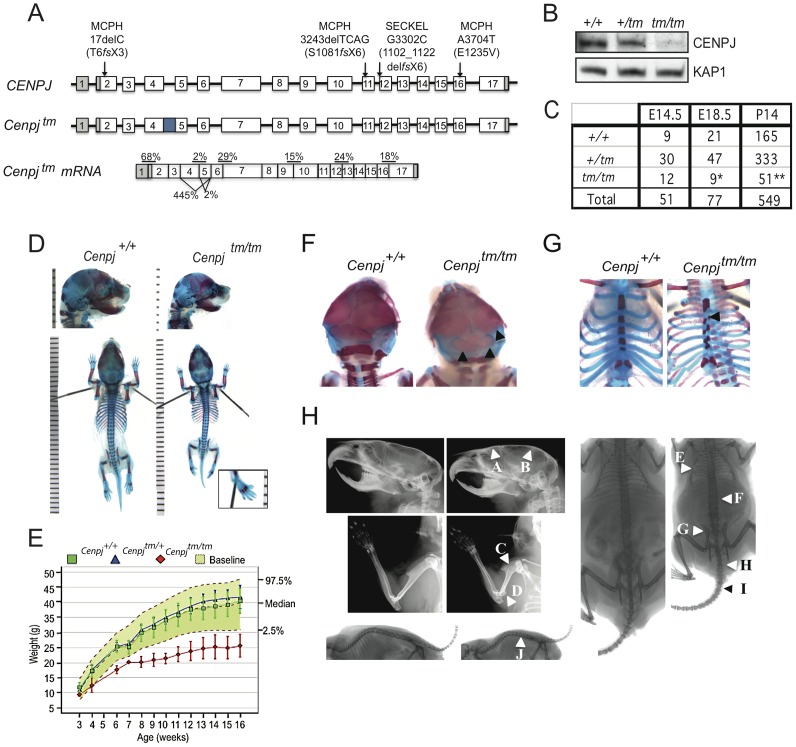
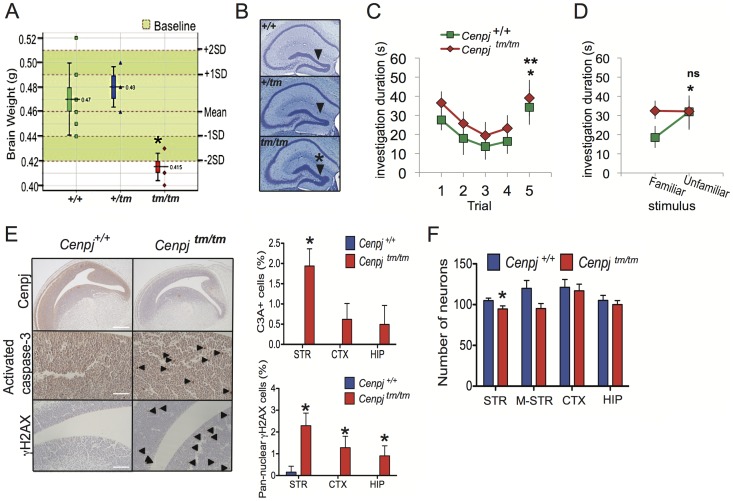
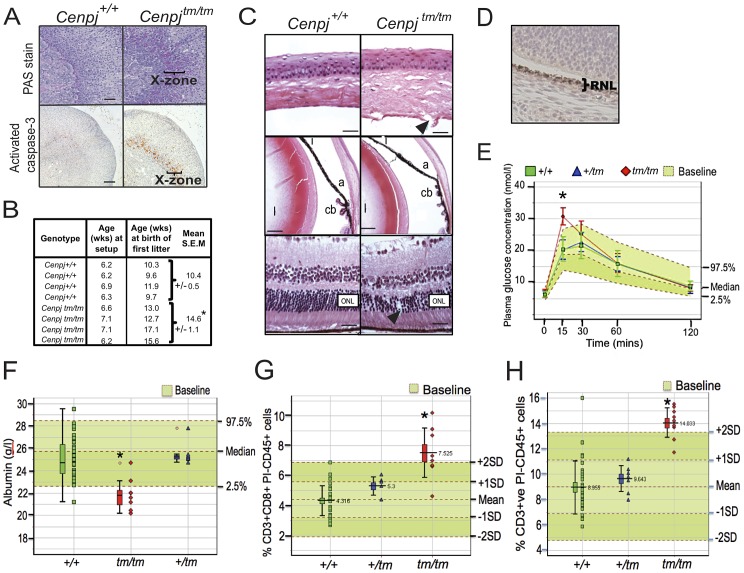
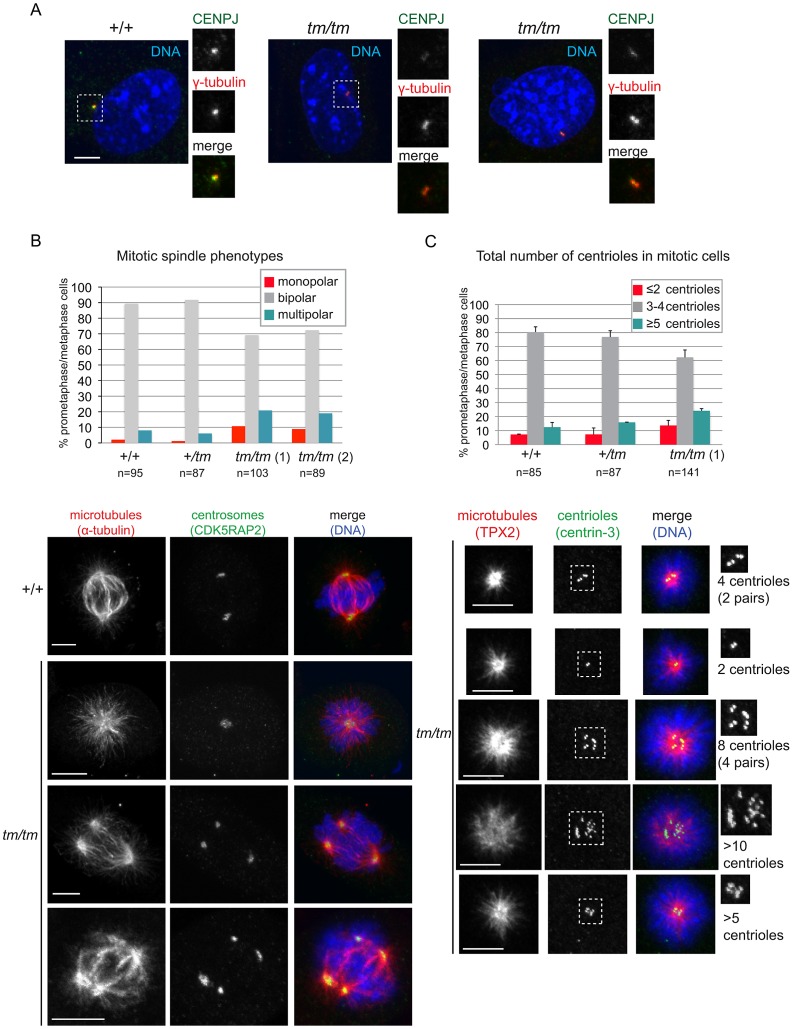
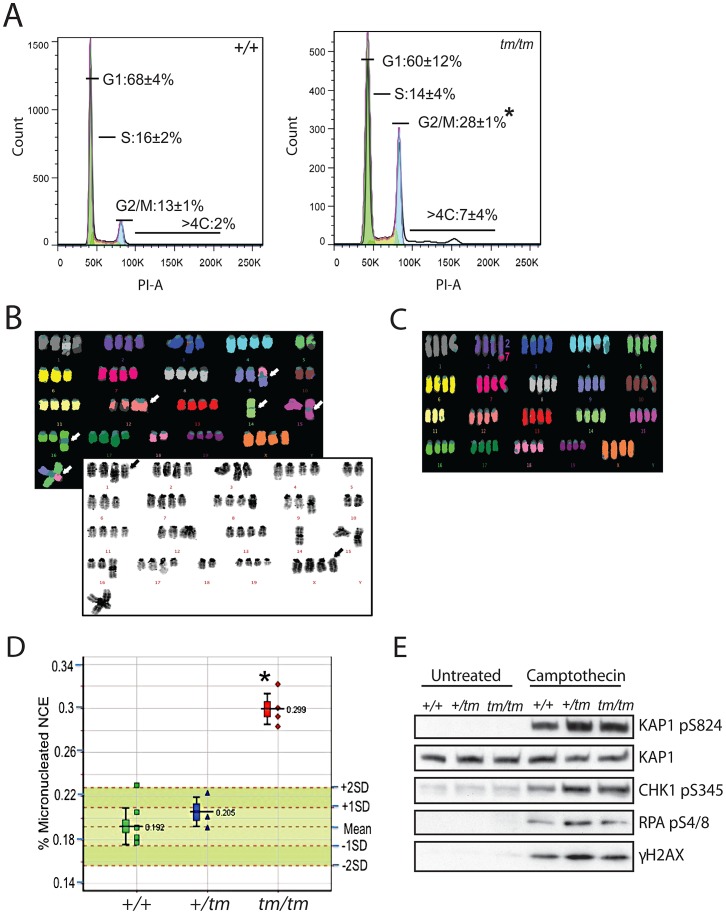
Disruption of the centromere protein J gene, CENPJ (CPAP, MCPH6, SCKL4), which is a highly conserved and ubiquitiously expressed centrosomal protein, has been associated with primary microcephaly and the microcephalic primordial dwarfism disorder Seckel syndrome. The mechanism by which disruption of CENPJ causes the proportionate, primordial growth failure that is characteristic of Seckel syndrome is unknown. By generating a hypomorphic allele of Cenpj, we have developed a mouse (Cenpj(tm/tm)) that recapitulates many of the clinical features of Seckel syndrome, including intrauterine dwarfism, microcephaly with memory impairment, ossification defects, and ocular and skeletal abnormalities, thus providing clear confirmation that specific mutations of CENPJ can cause Seckel syndrome. Immunohistochemistry revealed increased levels of DNA damage and apoptosis throughout Cenpj(tm/tm) embryos and adult mice showed an elevated frequency of micronucleus induction, suggesting that Cenpj-deficiency results in genomic instability. Notably, however, genomic instability was not the result of defective ATR-dependent DNA damage signaling, as is the case for the majority of genes associated with Seckel syndrome. Instead, Cenpj(tm/tm) embryonic fibroblasts exhibited irregular centriole and centrosome numbers and mono- and multipolar spindles, and many were near-tetraploid with numerical and structural chromosomal abnormalities when compared to passage-matched wild-type cells. Increased cell death due to mitotic failure during embryonic development is likely to contribute to the proportionate dwarfism that is associated with CENPJ-Seckel syndrome.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Hydranencephaly in CENPJ-related Seckel syndrome.Eur J Med Genet. 2022 Dec;65(12):104659. doi: 10.1016/j.ejmg.2022.104659. Epub 2022 Nov 2. Eur J Med Genet. 2022. PMID: 36334884
-
Analysis of centrosome and DNA damage response in PLK4 associated Seckel syndrome.Eur J Hum Genet. 2017 Oct;25(10):1118-1125. doi: 10.1038/ejhg.2017.120. Epub 2017 Aug 23. Eur J Hum Genet. 2017. PMID: 28832566 Free PMC article.
-
A novel leaky splice variant in centromere protein J (CENPJ)-associated Seckel syndrome.Ann Hum Genet. 2022 Sep;86(5):245-256. doi: 10.1111/ahg.12469. Epub 2022 Apr 22. Ann Hum Genet. 2022. PMID: 35451063
-
Consequences of Centrosome Dysfunction During Brain Development.Adv Exp Med Biol. 2017;1002:19-45. doi: 10.1007/978-3-319-57127-0_2. Adv Exp Med Biol. 2017. PMID: 28600781 Review.
-
Primary Autosomal Recessive Microcephalies and Seckel Syndrome Spectrum Disorders – RETIRED CHAPTER, FOR HISTORICAL REFERENCE ONLY.2009 Sep 1 [updated 2013 Oct 31]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. 2009 Sep 1 [updated 2013 Oct 31]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. PMID: 20301772 Free Books & Documents. Review.
Cited by
-
Molecular Genetics of Microcephaly Primary Hereditary: An Overview.Brain Sci. 2021 Apr 30;11(5):581. doi: 10.3390/brainsci11050581. Brain Sci. 2021. PMID: 33946187 Free PMC article. Review.
-
Cenpj Regulates Cilia Disassembly and Neurogenesis in the Developing Mouse Cortex.J Neurosci. 2019 Mar 13;39(11):1994-2010. doi: 10.1523/JNEUROSCI.1849-18.2018. Epub 2019 Jan 9. J Neurosci. 2019. PMID: 30626697 Free PMC article.
-
Pathway-specific effects of ADSL deficiency on neurodevelopment.Elife. 2022 Feb 8;11:e70518. doi: 10.7554/eLife.70518. Elife. 2022. PMID: 35133277 Free PMC article.
-
Centrobin-centrosomal protein 4.1-associated protein (CPAP) interaction promotes CPAP localization to the centrioles during centriole duplication.J Biol Chem. 2014 May 30;289(22):15166-78. doi: 10.1074/jbc.M113.531152. Epub 2014 Apr 3. J Biol Chem. 2014. PMID: 24700465 Free PMC article.
-
Mutations in PLK4, encoding a master regulator of centriole biogenesis, cause microcephaly, growth failure and retinopathy.Nat Genet. 2014 Dec;46(12):1283-1292. doi: 10.1038/ng.3122. Epub 2014 Oct 26. Nat Genet. 2014. PMID: 25344692 Free PMC article.
References
-
- Majewski F, Goecke T (1982) Studies of microcephalic primordial dwarfism I: approach to a delineation of the Seckel syndrome. American journal of medical genetics 12: 7–21. - PubMed
-
- Faivre L, Le Merrer M, Lyonnet S, Plauchu H, Dagoneau N, et al. (2002) Clinical and genetic heterogeneity of Seckel syndrome. American journal of medical genetics 112: 379–383. - PubMed
-
- Harsha Vardhan BG, Muthu MS, Saraswathi K, Koteeswaran D (2007) Bird-headed dwarf of Seckel. Journal of the Indian Society of Pedodontics and Preventive Dentistry 25 Suppl: S8–9. - PubMed
-
- Al-Dosari MS, Shaheen R, Colak D, Alkuraya FS (2010) Novel CENPJ mutation causes Seckel syndrome. J Med Genet 47: 411–414. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- 17043/CRUK_/Cancer Research UK/United Kingdom
- MC_UP_1502/1/MRC_/Medical Research Council/United Kingdom
- 12401/CRUK_/Cancer Research UK/United Kingdom
- NIH 1K08EY020530-01A1/EY/NEI NIH HHS/United States
- 098051/WT_/Wellcome Trust/United Kingdom
- 268536/ERC_/European Research Council/International
- G0901338/MRC_/Medical Research Council/United Kingdom
- 11224/CRUK_/Cancer Research UK/United Kingdom
- R01 EY018213/EY/NEI NIH HHS/United States
- 092096/WT_/Wellcome Trust/United Kingdom
- K08 EY020530/EY/NEI NIH HHS/United States
- 13031/CRUK_/Cancer Research UK/United Kingdom
- A11224/CRUK_/Cancer Research UK/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
