

Macroevolutionary dynamics and historical biogeography of primate diversification inferred from a species supermatrix
- PMID: 23166696
- PMCID: PMC3500307
- DOI: 10.1371/journal.pone.0049521
Macroevolutionary dynamics and historical biogeography of primate diversification inferred from a species supermatrix
Abstract
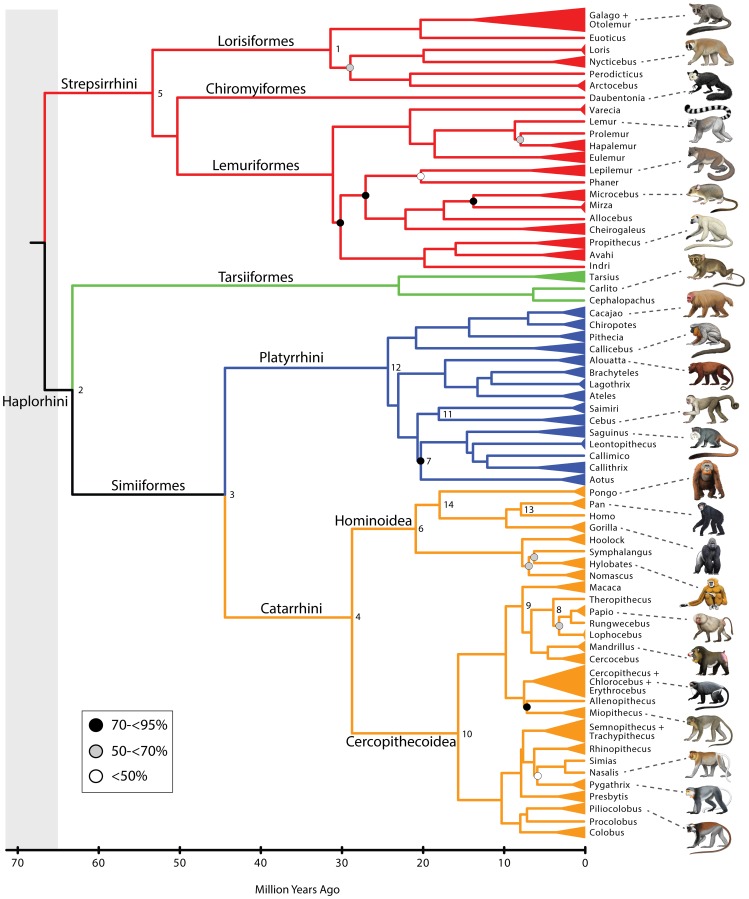
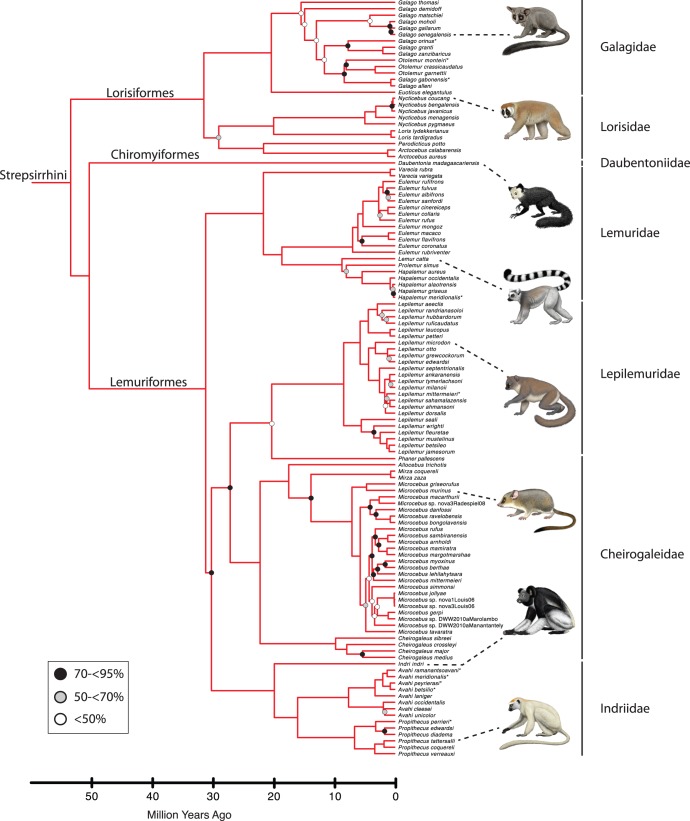
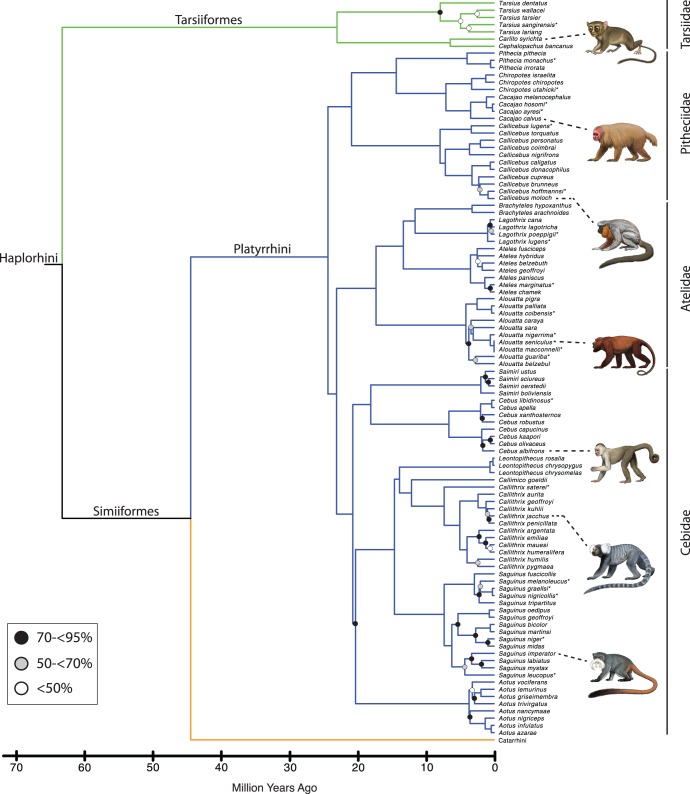
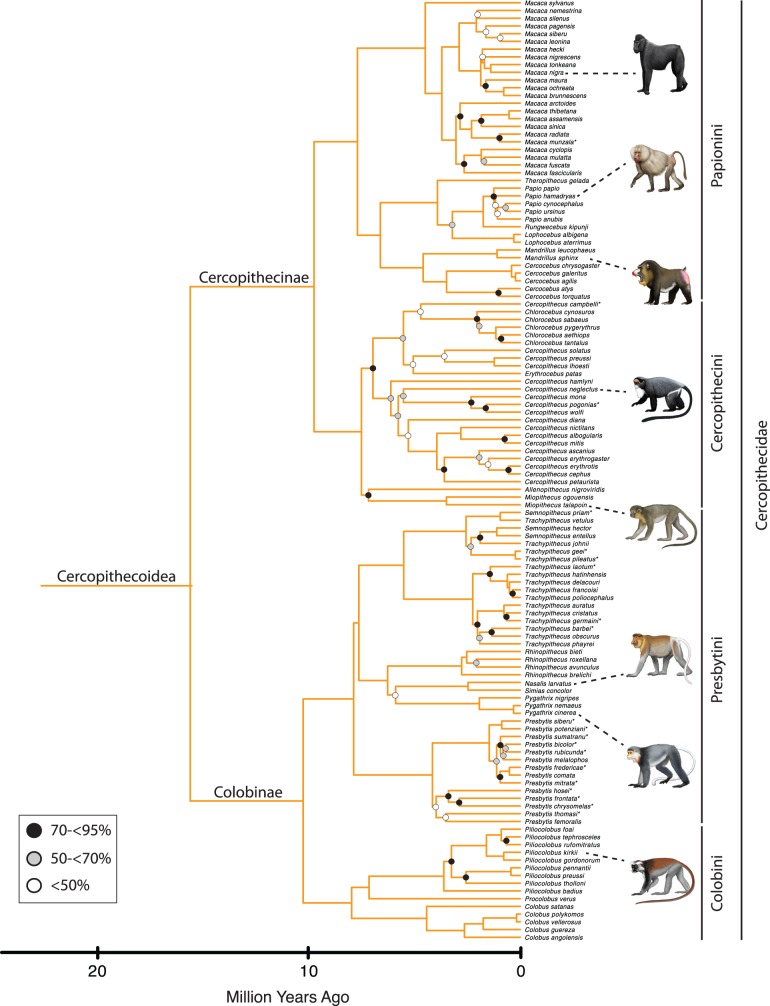
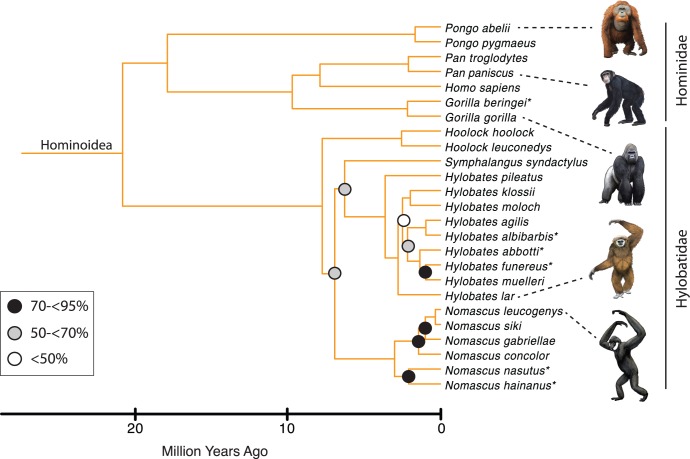
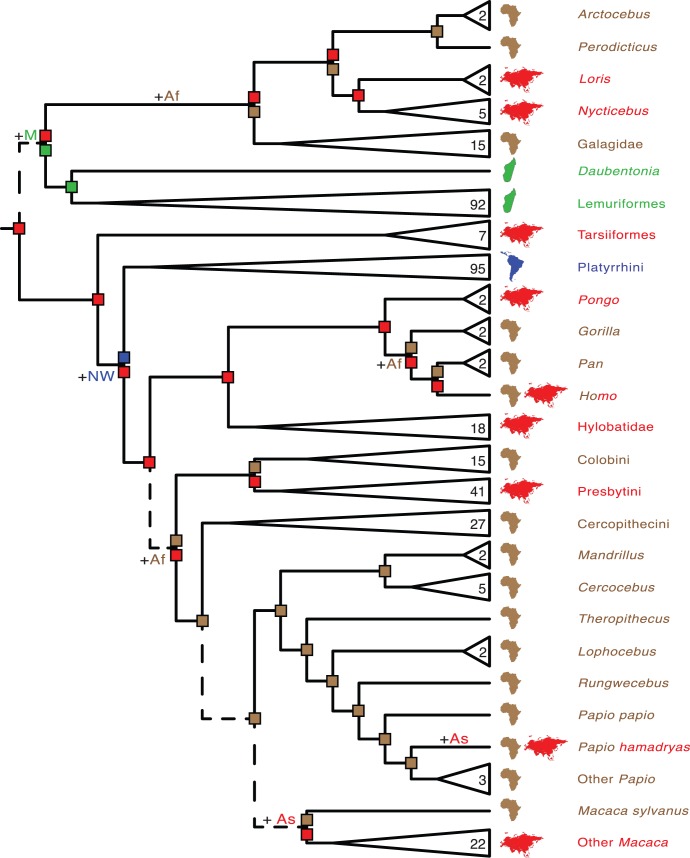
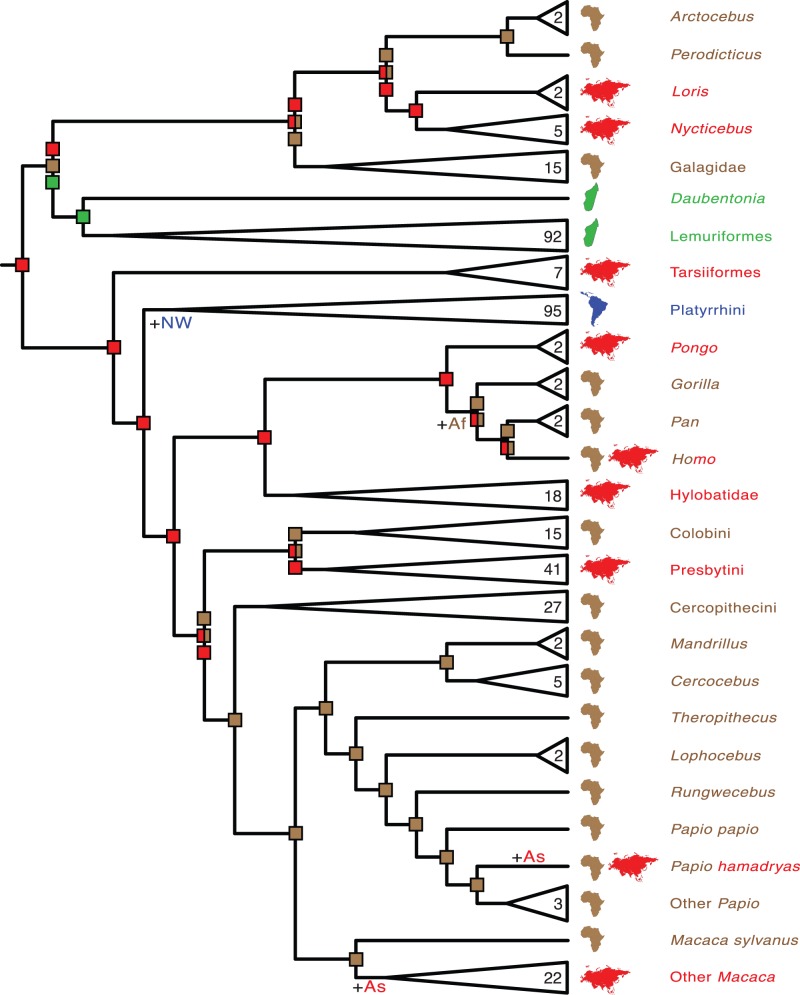
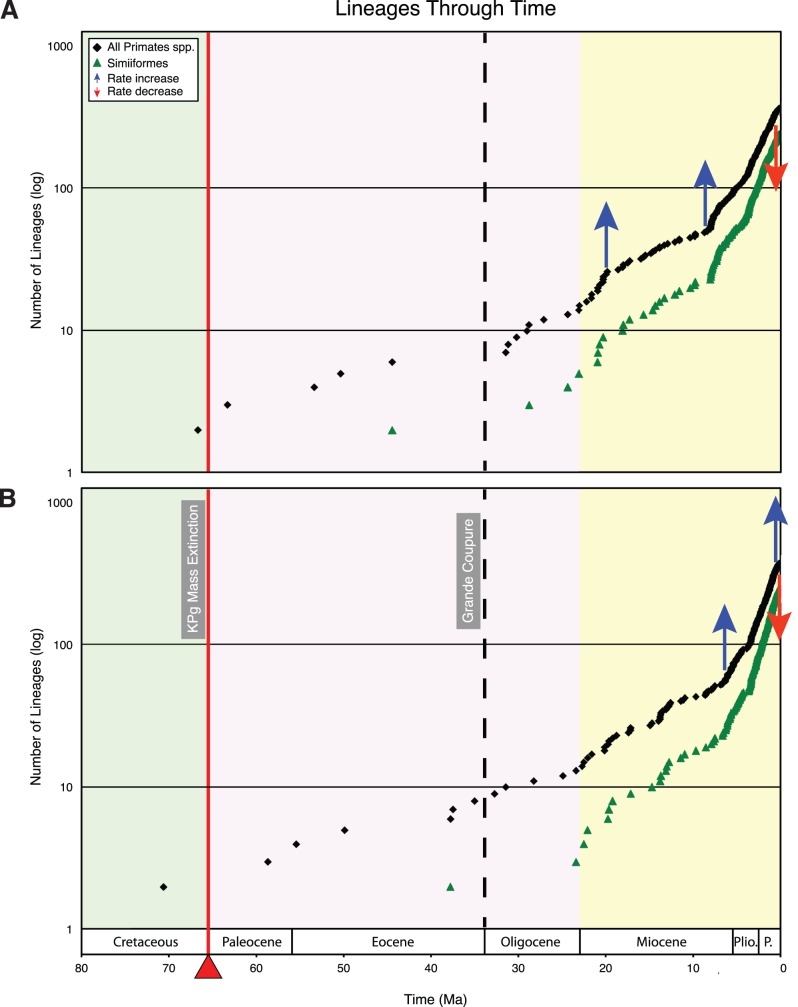
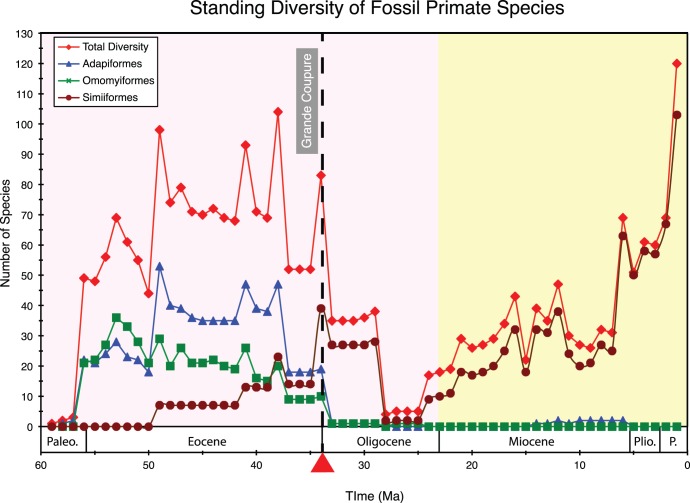
Phylogenetic relationships, divergence times, and patterns of biogeographic descent among primate species are both complex and contentious. Here, we generate a robust molecular phylogeny for 70 primate genera and 367 primate species based on a concatenation of 69 nuclear gene segments and ten mitochondrial gene sequences, most of which were extracted from GenBank. Relaxed clock analyses of divergence times with 14 fossil-calibrated nodes suggest that living Primates last shared a common ancestor 71-63 Ma, and that divergences within both Strepsirrhini and Haplorhini are entirely post-Cretaceous. These results are consistent with the hypothesis that the Cretaceous-Paleogene mass extinction of non-avian dinosaurs played an important role in the diversification of placental mammals. Previous queries into primate historical biogeography have suggested Africa, Asia, Europe, or North America as the ancestral area of crown primates, but were based on methods that were coopted from phylogeny reconstruction. By contrast, we analyzed our molecular phylogeny with two methods that were developed explicitly for ancestral area reconstruction, and find support for the hypothesis that the most recent common ancestor of living Primates resided in Asia. Analyses of primate macroevolutionary dynamics provide support for a diversification rate increase in the late Miocene, possibly in response to elevated global mean temperatures, and are consistent with the fossil record. By contrast, diversification analyses failed to detect evidence for rate-shift changes near the Eocene-Oligocene boundary even though the fossil record provides clear evidence for a major turnover event ("Grande Coupure") at this time. Our results highlight the power and limitations of inferring diversification dynamics from molecular phylogenies, as well as the sensitivity of diversification analyses to different species concepts.
Conflict of interest statement
Figures
References
-
- Silcox MT (2008) The biogeographic origins of Primates and Euprimates: east, west, north, or south of Eden? In: Sargis EJ, Dagosto M, eds. Mammalian evolutionary morphology: a tribute to Frederick S. Szalay. Dordrecht, The Netherlands: Springer. pp 199–231.
-
- Fabre PH, Rodrigues A, Douzery EJP (2009) Patterns of macroevolution among Primates inferred from a supermatrix of mitochondrial and nuclear DNA. Mol Phylogenet Evol 53: 808–825. - PubMed
-
- Groves CP (1993) Order Primates. In: Wilson DE, Reeder DM, eds. Mammal species of the world: a taxonomic and geographic reference, second edition. Washington and London: Smithsonian Institution Press. pp 243–277.
-
- Groves CP (2001) Primate taxonomy. Washington, DC: Smithsonian Institution Press. 350 p.
