

Modulation of antimicrobial host defense peptide gene expression by free fatty acids
- PMID: 23166711
- PMCID: PMC3499459
- DOI: 10.1371/journal.pone.0049558
Modulation of antimicrobial host defense peptide gene expression by free fatty acids
Abstract
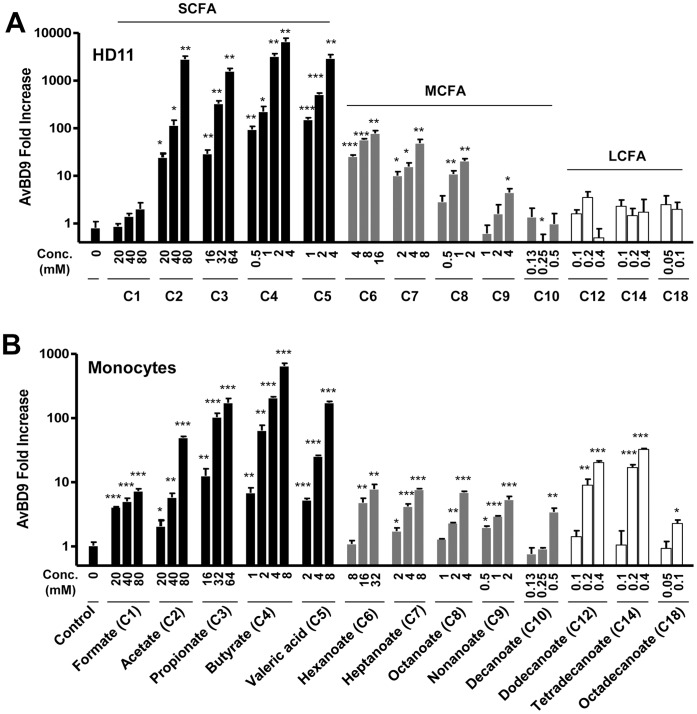
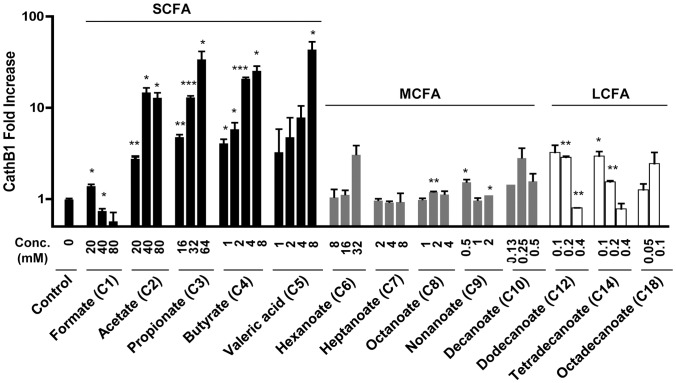
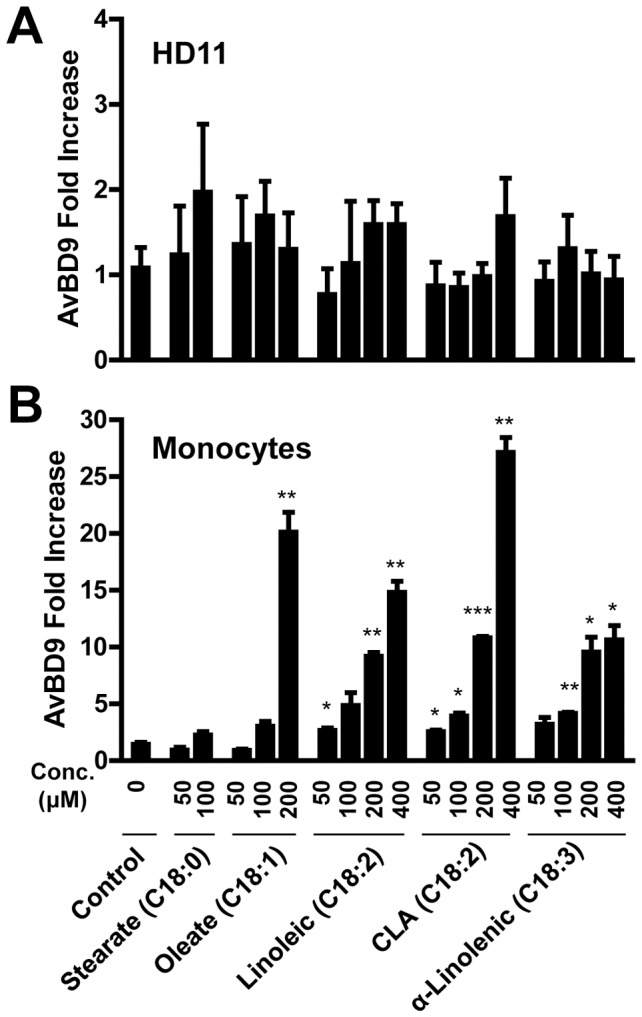
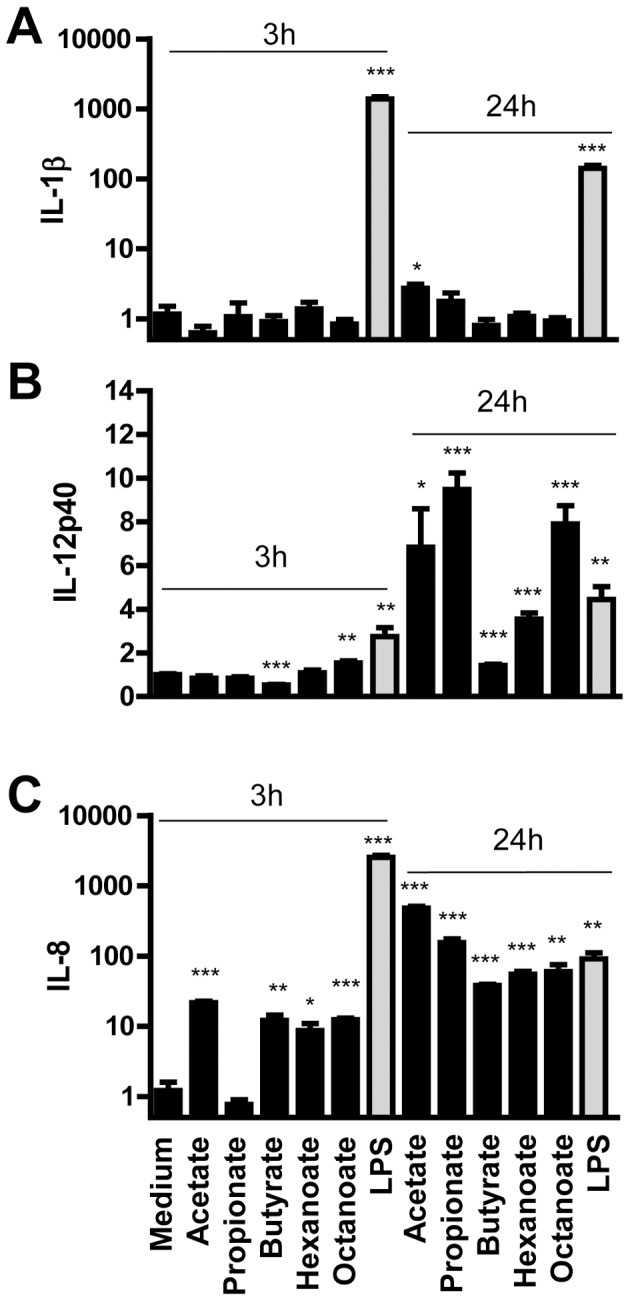
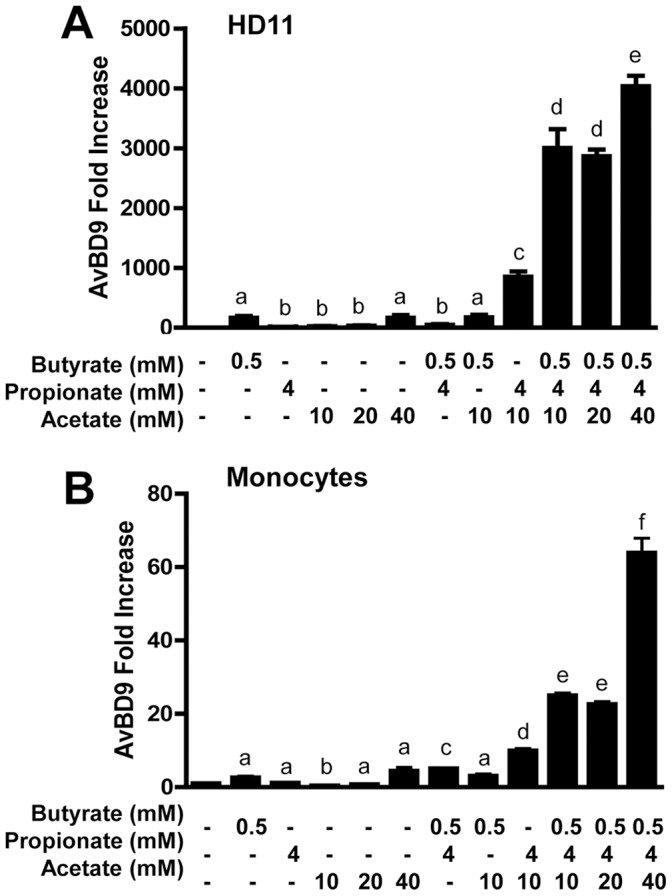
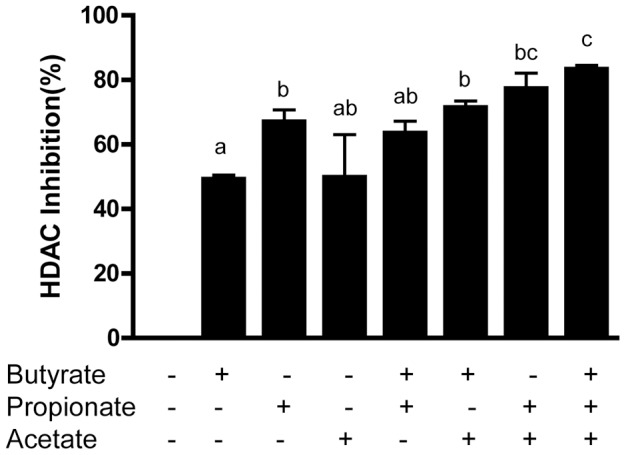
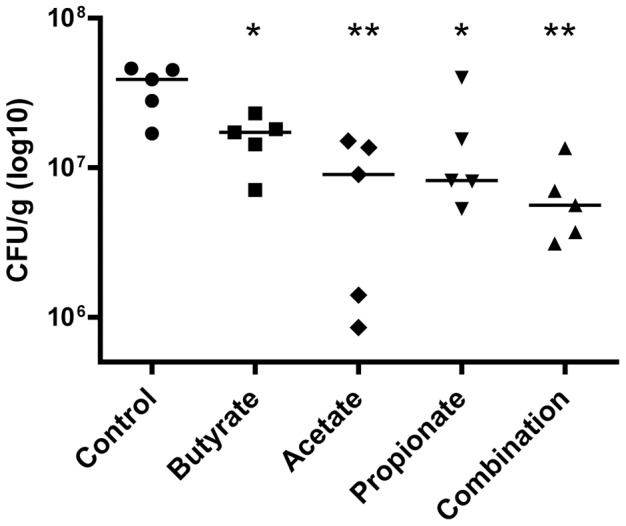
Routine use of antibiotics at subtherapeutic levels in animal feed drives the emergence of antimicrobial resistance. Development of antibiotic-alternative approaches to disease control and prevention for food animals is imperatively needed. Previously, we showed that butyrate, a major species of short-chain fatty acids (SCFAs) fermented from undigested fiber by intestinal microflora, is a potent inducer of endogenous antimicrobial host defense peptide (HDP) genes in the chicken (PLoS One 2011, 6: e27225). In the present study, we further revealed that, in chicken HD11 macrophages and primary monocytes, induction of HDPs is largely in an inverse correlation with the aliphatic hydrocarbon chain length of free fatty acids, with SCFAs being the most potent, medium-chain fatty acids moderate and long-chain fatty acids marginal. Additionally, three SCFAs, namely acetate, propionate, and butyrate, exerted a strong synergy in augmenting HDP gene expression in chicken cells. Consistently, supplementation of chickens with a combination of three SCFAs in water resulted in a further reduction of Salmonella enteritidis in the cecum as compared to feeding of individual SCFAs. More importantly, free fatty acids enhanced HDP gene expression without triggering proinflammatory interleukin-1β production. Taken together, oral supplementation of SCFAs is capable of boosting host immunity and disease resistance, with potential for infectious disease control and prevention in animal agriculture without relying on antibiotics.
Conflict of interest statement
Figures
Similar articles
-
Induction of porcine host defense peptide gene expression by short-chain fatty acids and their analogs.PLoS One. 2013 Aug 30;8(8):e72922. doi: 10.1371/journal.pone.0072922. eCollection 2013. PLoS One. 2013. PMID: 24023657 Free PMC article.
-
Butyrate enhances disease resistance of chickens by inducing antimicrobial host defense peptide gene expression.PLoS One. 2011;6(11):e27225. doi: 10.1371/journal.pone.0027225. Epub 2011 Nov 4. PLoS One. 2011. PMID: 22073293 Free PMC article.
-
Differential regulation of human cathelicidin LL-37 by free fatty acids and their analogs.Peptides. 2013 Dec;50:129-38. doi: 10.1016/j.peptides.2013.10.008. Epub 2013 Oct 18. Peptides. 2013. PMID: 24140860
-
Microbiota-derived short-chain fatty acids and modulation of host-derived peptides formation: Focused on host defense peptides.Biomed Pharmacother. 2023 Jun;162:114586. doi: 10.1016/j.biopha.2023.114586. Epub 2023 Mar 28. Biomed Pharmacother. 2023. PMID: 36989711 Review.
-
Nutritional Modulation of Host Defense Peptide Synthesis: A Novel Host-Directed Antimicrobial Therapeutic Strategy?Adv Nutr. 2024 Sep;15(9):100277. doi: 10.1016/j.advnut.2024.100277. Epub 2024 Jul 23. Adv Nutr. 2024. PMID: 39053604 Free PMC article. Review.
Cited by
-
Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health.Gut Microbes. 2020 Nov 9;12(1):1802866. doi: 10.1080/19490976.2020.1802866. Gut Microbes. 2020. PMID: 32835590 Free PMC article. Review.
-
The Microbial Pecking Order: Utilization of Intestinal Microbiota for Poultry Health.Microorganisms. 2019 Sep 20;7(10):376. doi: 10.3390/microorganisms7100376. Microorganisms. 2019. PMID: 31547217 Free PMC article. Review.
-
Regulation of bacterial pathogenesis by intestinal short-chain Fatty acids.Adv Appl Microbiol. 2013;85:93-118. doi: 10.1016/B978-0-12-407672-3.00003-4. Adv Appl Microbiol. 2013. PMID: 23942149 Free PMC article. Review.
-
Induction of porcine host defense peptide gene expression by short-chain fatty acids and their analogs.PLoS One. 2013 Aug 30;8(8):e72922. doi: 10.1371/journal.pone.0072922. eCollection 2013. PLoS One. 2013. PMID: 24023657 Free PMC article.
-
Effects of Microencapsulated Blends of Organics Acids (OA) and Essential Oils (EO) as a Feed Additive for Broiler Chicken. A Focus on Growth Performance, Gut Morphology and Microbiology.Animals (Basel). 2020 Mar 6;10(3):442. doi: 10.3390/ani10030442. Animals (Basel). 2020. PMID: 32155791 Free PMC article.
References
-
- Zasloff M (2002) Antimicrobial peptides of multicellular organisms. Nature 415: 389–395. - PubMed
-
- Brogden KA, Ackermann M, McCray PB Jr, Tack BF (2003) Antimicrobial peptides in animals and their role in host defences. Int J Antimicrob Agents 22: 465–478. - PubMed
-
- Hancock RE, Sahl HG (2006) Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol 24: 1551–1557. - PubMed
-
- Yang D, Biragyn A, Hoover DM, Lubkowski J, Oppenheim JJ (2004) Multiple roles of antimicrobial defensins, cathelicidins, and eosinophil-derived neurotoxin in host defense. Annu Rev Immunol 22: 181–215. - PubMed
-
- Van Immerseel F, Russell JB, Flythe MD, Gantois I, Timbermont L, et al. (2006) The use of organic acids to combat Salmonella in poultry: a mechanistic explanation of the efficacy. Avian Pathol 35: 182–188. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
