

The β-catenin destruction complex
- PMID: 23169527
- PMCID: PMC3579403
- DOI: 10.1101/cshperspect.a007898
The β-catenin destruction complex
Abstract
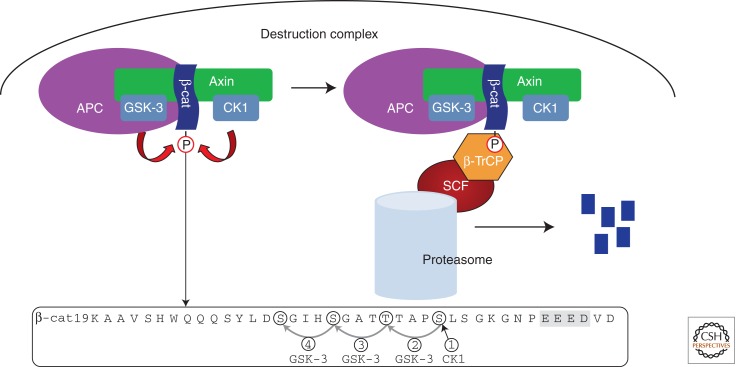
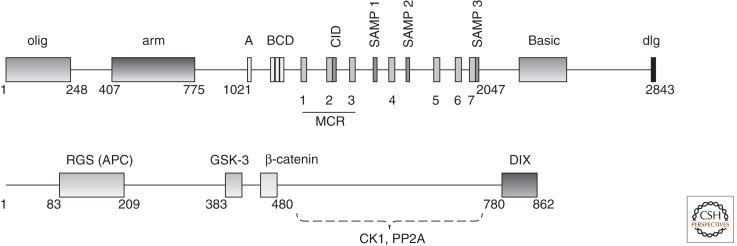
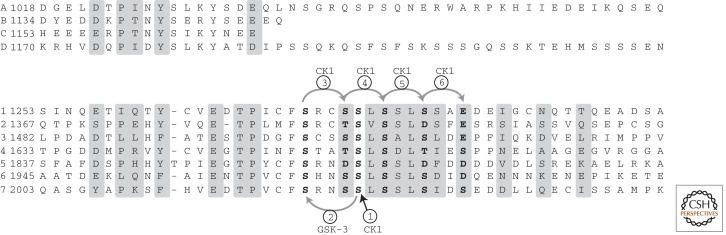
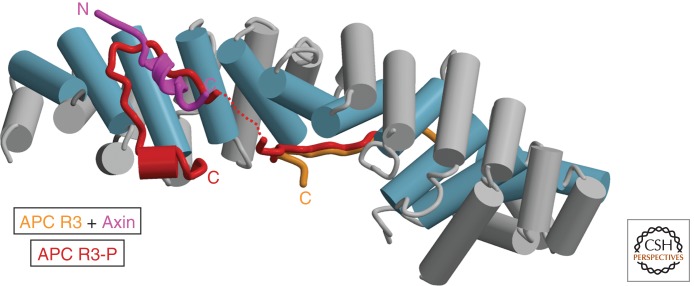
The Wnt/β-catenin pathway is highly regulated to insure the correct temporal and spatial activation of its target genes. In the absence of a Wnt stimulus, the transcriptional coactivator β-catenin is degraded by a multiprotein "destruction complex" that includes the tumor suppressors Axin and adenomatous polyposis coli (APC), the Ser/Thr kinases GSK-3 and CK1, protein phosphatase 2A (PP2A), and the E3-ubiquitin ligase β-TrCP. The complex generates a β-TrCP recognition site by phosphorylation of a conserved Ser/Thr-rich sequence near the β-catenin amino terminus, a process that requires scaffolding of the kinases and β-catenin by Axin. Ubiquitinated β-catenin is degraded by the proteasome. The molecular mechanisms that underlie several aspects of destruction complex function are poorly understood, particularly the role of APC. Here we review the molecular mechanisms of destruction complex function and discuss several potential roles of APC in β-catenin destruction.
Figures
References
-
- Albuquerque C, Breukel C, van der Luijt R, Fidalgo P, Lage P, Slors FJ, Leitao CN, Fodde R, Smits R 2002. The “just-right” signaling model: APC somatic mutations are selected based on a specific level of activation of the β-catenin signaling cascade. Hum Mol Genet 11: 1549–1560 - PubMed
-
- Behrens J, von Kries JP, Kühl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W 1996. Functional interaction of β-catenin with the transcription factor LEF-1. Nature 382: 638–642 - PubMed
-
- Behrens J, Jerchow B-A, Würtele M, Grimm J, Asbrand C, Wirtz R, Kühl M, Wedlich D, Birchmeier W 1998. Functional interaction of an Axin homolog, conductin, with β-catenin, APC, and GSK3β. Science 280: 596–599 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials