

The development and application of new crystallization method for tobacco mosaic virus coat protein
- PMID: 23171808
- PMCID: PMC3560112
- DOI: 10.1186/1743-422X-9-279
The development and application of new crystallization method for tobacco mosaic virus coat protein
Abstract
Background: Although tobacco mosaic virus (TMV) coat protein (CP) has been isolated from virus particles and its crystals have grown in ammonium sulfate buffers for many years, to date, no one has reported on the crystallization of recombinant TMV-CP connecting peptides expressed in E. coli.
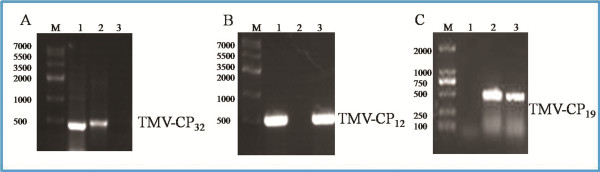
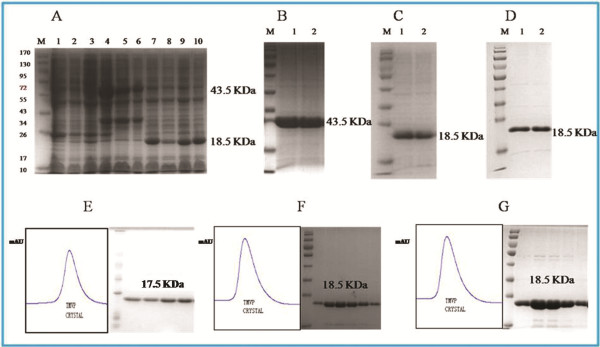
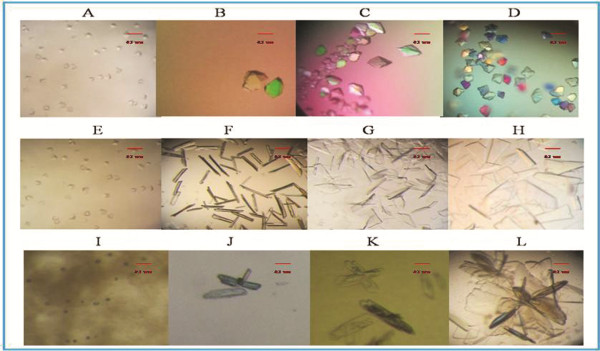
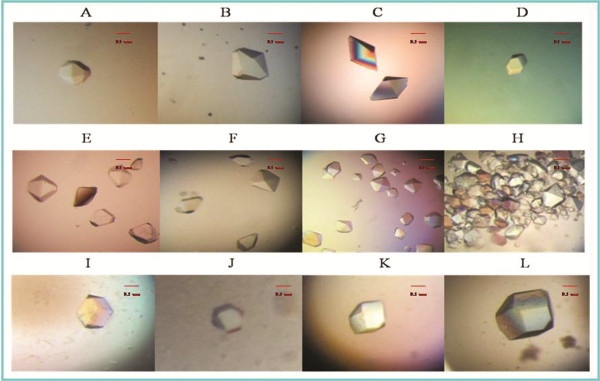
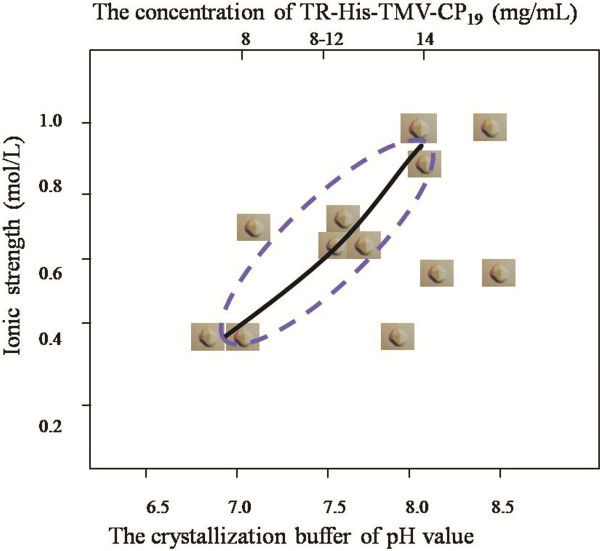
Methods: In the present papers genetically engineered TMV-CP was expressed, into which hexahistidine (His) tags or glutathione-S-transferase (GST) tags were incorporated. Considering that GST-tags are long peptides and His-tags are short peptides, an attempt was made to grow crystals of TMV-CP cleaved GST-tags (WT-TMV-CP32) and TMV-CP incorporated His-tags (WT-His-TMV-CP12) simultaneously in ammonium sulfate buffers and commercial crystallization reagents. It was found that the 20S disk form of WT-TMV-CP32 and WT-His-TMV-CP12 did not form high resolution crystals by using various crystallization buffers and commercial crystallization reagents. Subsequently, a new experimental method was adopted in which a range of truncated TMV-CP was constructed by removing several amino acids from the N- or the C-terminal, and high resolution crystals were grown in ammonium sulfate buffers and commercial crystallization reagents.
Results: The new crystallization method was developed and 3.0 Å resolution macromolecular crystal was thereby obtained by removing four amino acids at the C-terminal of His-TMV-CP and connecting six His-tags at the N-terminal of His-TMV-CP (TR-His-TMV-CP19). The Four-layer aggregate disk structure of TR-His-TMV-CP19 was solved. This phenomenon showed that peptides at the C-terminus hindered the growth of high resolution crystals and the peptides interactions at the N-terminus were attributed to the quality of TMV-CP crystals.
Conclusion: A 3.0 Å resolution macromolecular crystal of TR-His-TMV-CP19 was obtained and the corresponding structure was solved by removing four amino acids at the C-terminus of TMV-CP and connecting His-tags at the N-terminus of TMV-CP. It indicated that short peptides influenced the resolution of TMV-CP crystals.
Figures


Similar articles
-
Crystal structure of a four-layer aggregate of engineered TMV CP implies the importance of terminal residues for oligomer assembly.PLoS One. 2013 Nov 4;8(11):e77717. doi: 10.1371/journal.pone.0077717. eCollection 2013. PLoS One. 2013. PMID: 24223721 Free PMC article.
-
Role of hexahistidine in directed nanoassemblies of tobacco mosaic virus coat protein.ACS Nano. 2011 Mar 22;5(3):1606-16. doi: 10.1021/nn1025719. Epub 2011 Feb 25. ACS Nano. 2011. PMID: 21361370
-
In vitro assembly of Tobacco mosaic virus coat protein variants derived from fission yeast expression clones or plants.J Virol Methods. 2010 Jun;166(1-2):77-85. doi: 10.1016/j.jviromet.2010.02.026. Epub 2010 Feb 26. J Virol Methods. 2010. PMID: 20219539
-
Coat-protein-mediated resistance to tobacco mosaic virus: discovery mechanisms and exploitation.Philos Trans R Soc Lond B Biol Sci. 1999 Mar 29;354(1383):659-64. doi: 10.1098/rstb.1999.0418. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10212946 Free PMC article. Review.
-
Molecular assembly of tobacco mosaic virus in vitro.Adv Biophys. 1986;22:95-149. doi: 10.1016/0065-227x(86)90004-3. Adv Biophys. 1986. PMID: 3551520 Review.
Cited by
-
Extensive libraries of gene truncation variants generated by in vitro transposition.Nucleic Acids Res. 2017 Jun 2;45(10):e78. doi: 10.1093/nar/gkx030. Nucleic Acids Res. 2017. PMID: 28130425 Free PMC article.
-
Chemical and Biological Investigations of Antiviral Agents Against Plant Viruses Conducted in China in the 21st Century.Genes (Basel). 2024 Dec 23;15(12):1654. doi: 10.3390/genes15121654. Genes (Basel). 2024. PMID: 39766921 Free PMC article. Review.
-
Design, Synthesis, and Antiviral and Fungicidal Activities of 4-Oxo-4H-quinolin-1-yl Acylhydrazone Derivatives.ACS Omega. 2024 Aug 14;9(34):36671-36681. doi: 10.1021/acsomega.4c05046. eCollection 2024 Aug 27. ACS Omega. 2024. PMID: 39220544 Free PMC article.
-
Chalcone derivatives containing 1,2,4-triazole and pyridine moiety: design, synthesis, and antiviral activity.Mol Divers. 2024 Dec 2. doi: 10.1007/s11030-024-11049-7. Online ahead of print. Mol Divers. 2024. PMID: 39617870
-
Crystal structure of a four-layer aggregate of engineered TMV CP implies the importance of terminal residues for oligomer assembly.PLoS One. 2013 Nov 4;8(11):e77717. doi: 10.1371/journal.pone.0077717. eCollection 2013. PLoS One. 2013. PMID: 24223721 Free PMC article.
References
-
- Caspar DLD. Assembly and stability of the tobacco mosaic virus particle. Adv Protein Chem. 1963;18:37–121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
