

Cardiolipin-dependent reconstitution of respiratory supercomplexes from purified Saccharomyces cerevisiae complexes III and IV
- PMID: 23172229
- PMCID: PMC3537037
- DOI: 10.1074/jbc.M112.425876
Cardiolipin-dependent reconstitution of respiratory supercomplexes from purified Saccharomyces cerevisiae complexes III and IV
Abstract
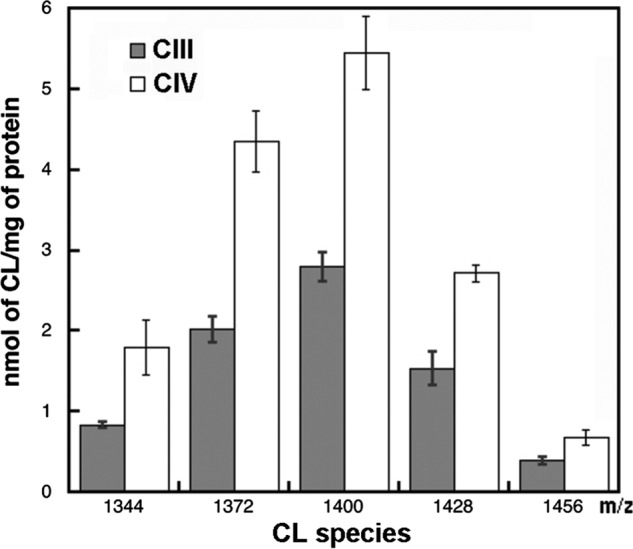
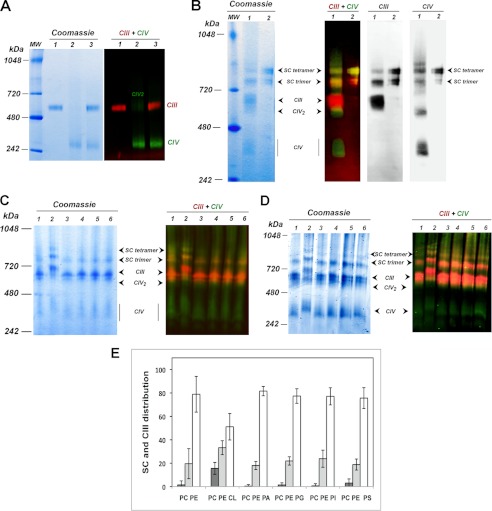
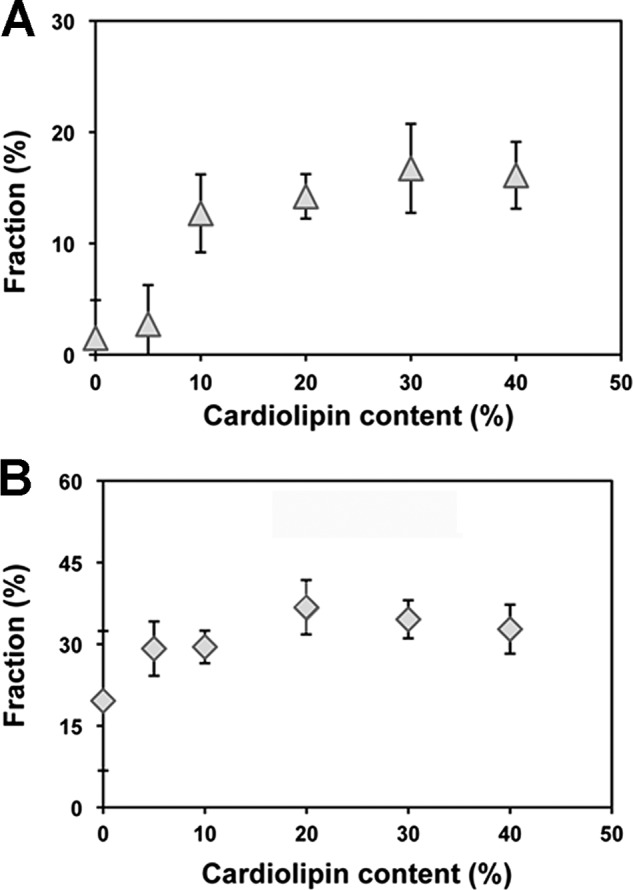
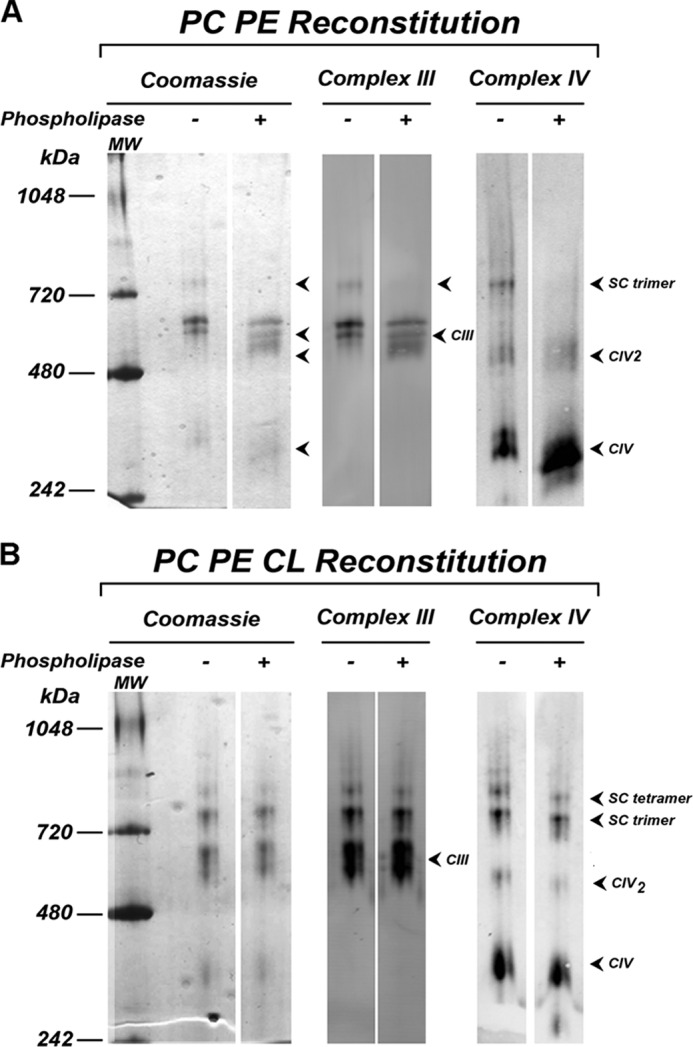
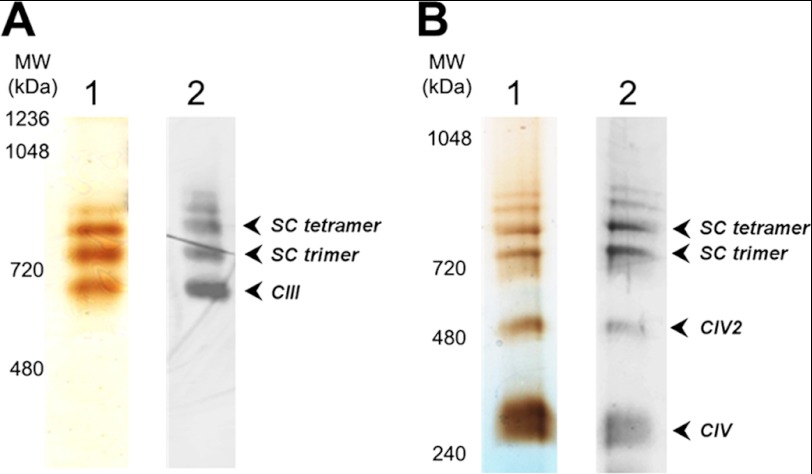
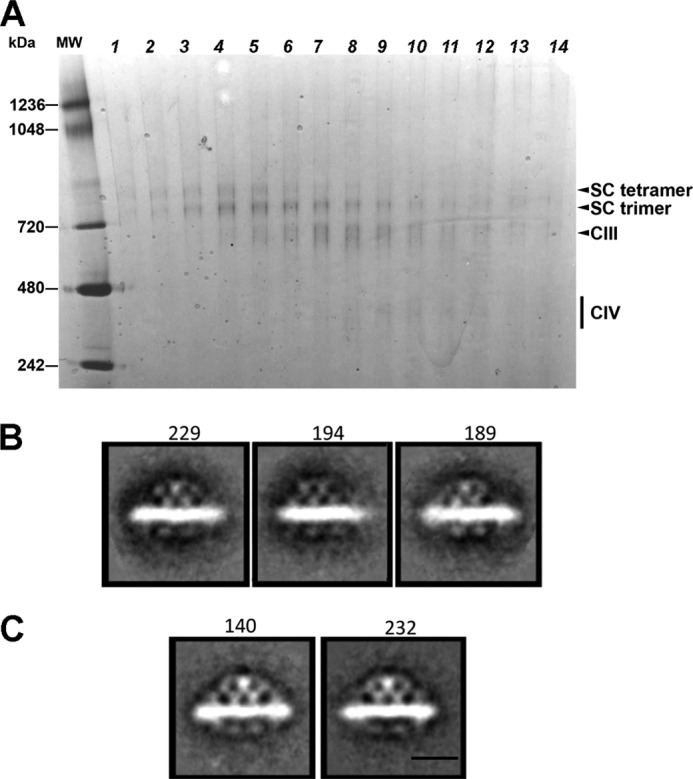
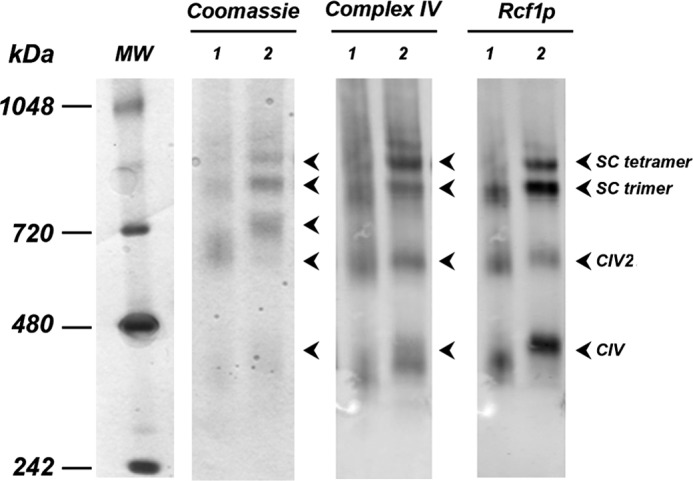
Here, we report for the first time in vitro reconstitution of the respiratory supercomplexes from individual complexes III and IV. Complexes III and IV were purified from Saccharomyces cerevisiae mitochondria. Complex III contained eight molecules of cardiolipin, and complex IV contained two molecules of cardiolipin, as determined by electrospray ionization-mass spectrometry. Complex IV also contained Rcf1p. No supercomplexes were formed upon mixing of the purified complexes, and low amounts of the supercomplex trimer III(2)IV(1) were formed after reconstitution into proteoliposomes containing only phosphatidylcholine and phosphatidylethanolamine. Further addition of cardiolipin to the proteoliposome reconstitution mixture resulted in distinct formation of both the III(2)IV(1) supercomplex trimer and III(2)IV(2) supercomplex tetramer. No other anionic phospholipid was as effective as cardiolipin in supporting tetramer formation. Phospholipase treatment of complex IV prevented trimer formation in the absence of cardiolipin. Both trimer and tetramer formations were restored by cardiolipin. Analysis of the reconstituted tetramer by single particle electron microscopy confirmed native organization of individual complexes within the supercomplex. In conclusion, although some trimer formation occurred dependent only on tightly bound cardiolipin, tetramer formation required additional cardiolipin. This is consistent with the high cardiolipin content in the native tetramer. The dependence on cardiolipin for supercomplex formation suggests that changes in cardiolipin levels resulting from changes in physiological conditions may control the equilibrium between individual respiratory complexes and supercomplexes in vivo.
Figures
References
-
- Chance B., Williams G. R. (1955) A method for the localization of sites for oxidative phosphorylation. Nature 176, 250–254 - PubMed
-
- Dudkina N. V., Kouril R., Peters K., Braun H. P., Boekema E. J. (2010) Structure and function of mitochondrial supercomplexes. Biochim. Biophys. Acta 1797, 664–670 - PubMed
-
- Lenaz G., Genova M. L. (2012) Supramolecular organisation of the mitochondrial respiratory chain. A new challenge for the mechanism and control of oxidative phosphorylation. Adv. Exp. Med. Biol. 748, 107–144 - PubMed
-
- Schägger H. (2001) Respiratory chain supercomplexes. IUBMB Life 52, 119–128 - PubMed
-
- Stuart R. A. (2008) Supercomplex organization of the oxidative phosphorylation enzymes in yeast mitochondria. J. Bioenerg. Biomembr. 40, 411–417 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
