

Racemose inflorescences of monocots: structural and morphogenetic interaction at the flower/inflorescence level
- PMID: 23172413
- PMCID: PMC3828938
- DOI: 10.1093/aob/mcs246
Racemose inflorescences of monocots: structural and morphogenetic interaction at the flower/inflorescence level
Abstract
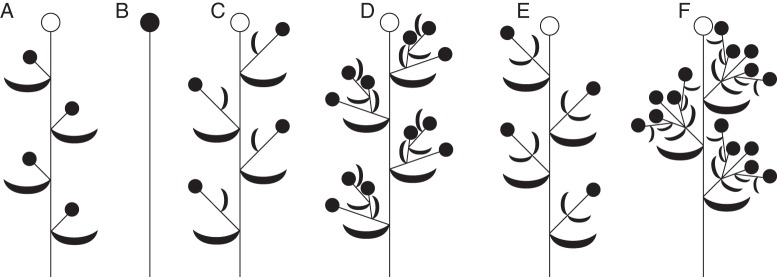
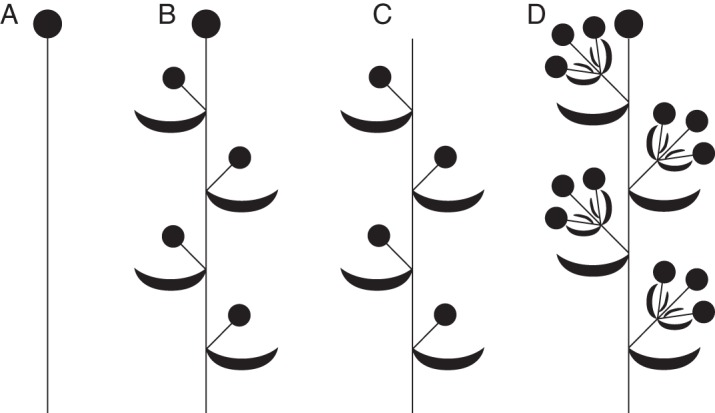
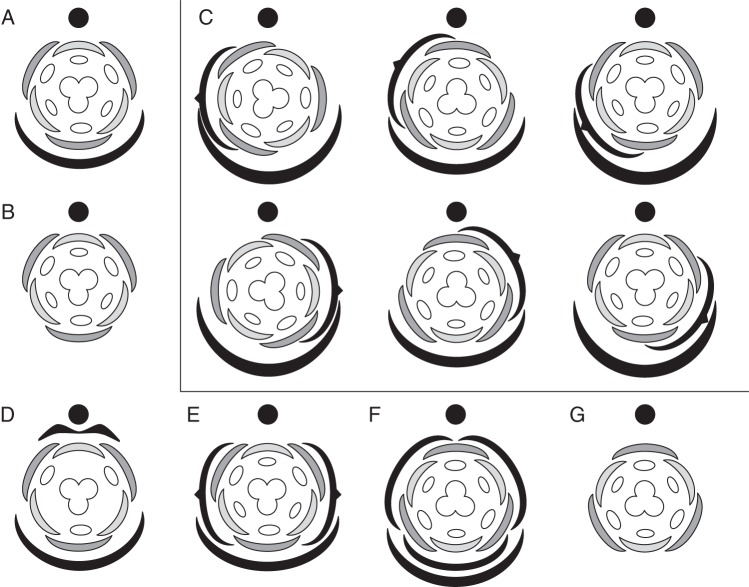
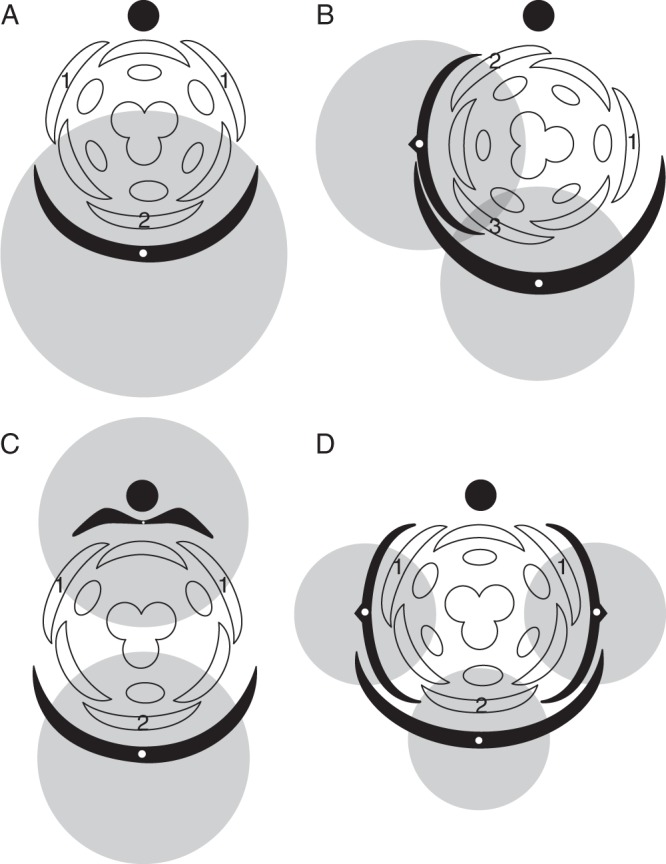
Background: Understanding and modelling early events of floral meristem patterning and floral development requires consideration of positional information regarding the organs surrounding the floral meristem, such as the flower-subtending bracts (FSBs) and floral prophylls (bracteoles). In common with models of regulation of floral patterning, the simplest models of phyllotaxy consider only unbranched uniaxial systems. Racemose inflorescences and thyrses offer a useful model system for investigating morphogenetic interactions between organs belonging to different axes.
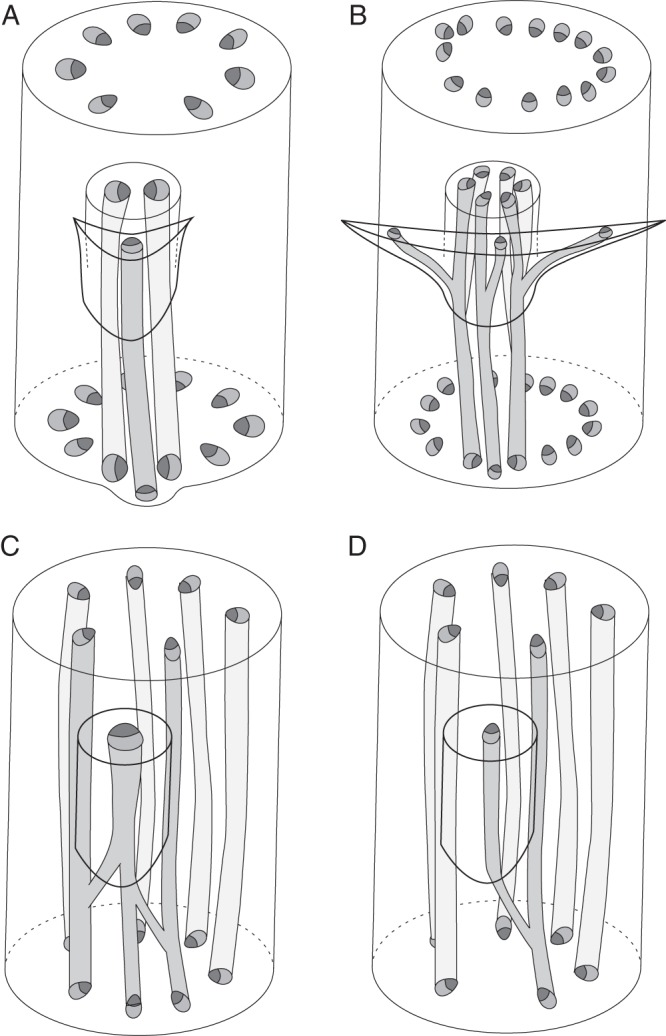
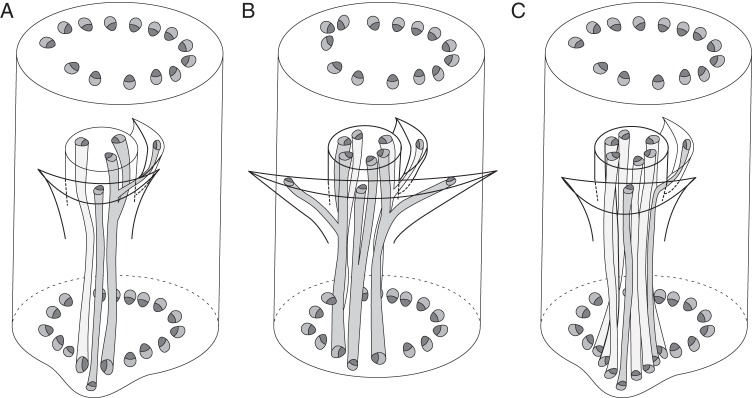
Scope: This review considers (1) racemose inflorescences of early-divergent and lilioid monocots and their possible relationship with other inflorescence types, (2) hypotheses on the morphogenetic significance of phyllomes surrounding developing flowers, (3) patterns of FSB reduction and (4) vascular patterns in the primary inflorescence axis and lateral pedicels.
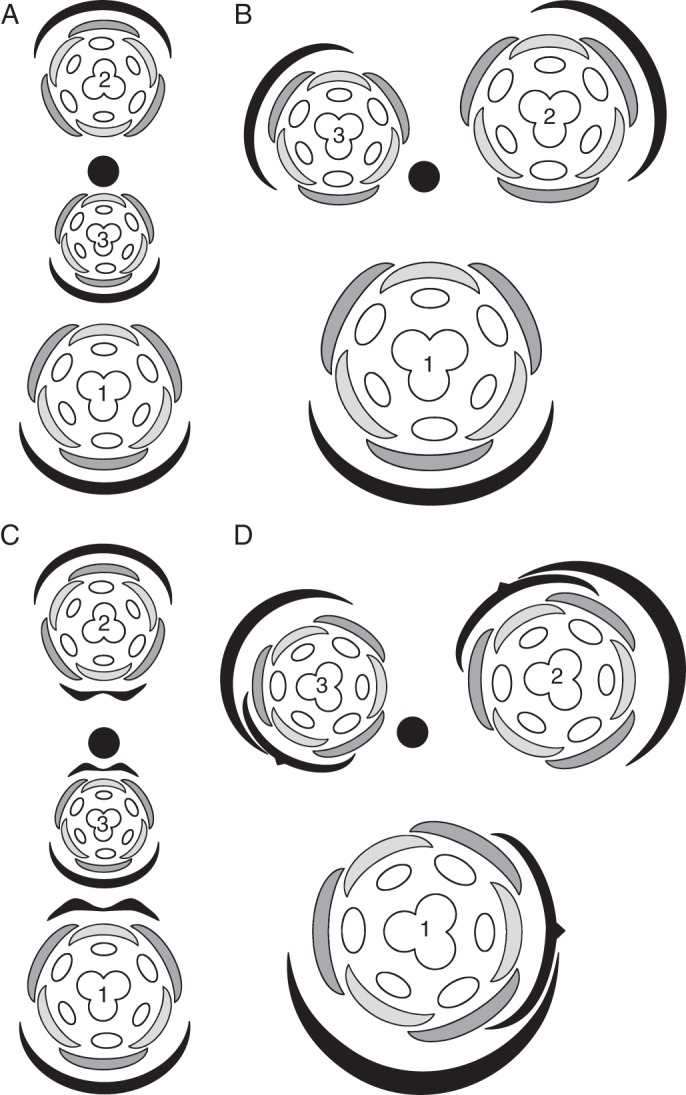
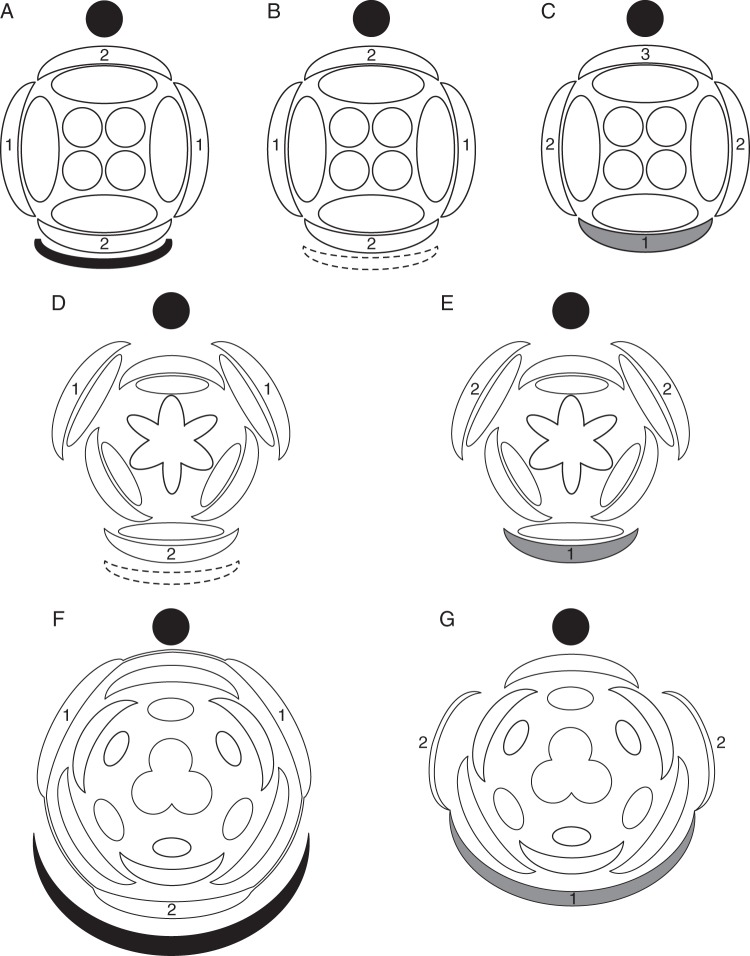
Conclusions: Racemose (partial) inflorescences represent the plesiomorphic condition in monocots. The presence or absence of a terminal flower or flower-like structure is labile among early-divergent monocots. In some Alismatales, a few-flowered racemose inflorescence can be entirely transformed into a terminal 'flower'. The presence or absence and position of additional phyllomes on the lateral pedicels represent important taxonomic markers and key features in regulation of flower patterning. Racemose inflorescences with a single floral prophyll are closely related to thyrses. Floral patterning is either unidirectional or simultaneous in species that lack a floral prophyll or possess a single adaxial floral prophyll and usually spiral in the outer perianth whorl in species with a transversely oriented floral prophyll. Inhibitory fields of surrounding phyllomes are relevant but insufficient to explain these patterns; other important factors are meristem space economy and/or the inhibitory activity of the primary inflorescence axis. Two patterns of FSB reduction exist in basal monocots: (1) complete FSB suppression (cryptic flower-subtending bract) and (2) formation of a 'hybrid' organ by overlap of the developmental programmes of the FSB and the first abaxial organ formed on the floral pedicel. FSB reduction affects patterns of interaction between the conductive systems of the flower and the primary inflorescence axis.
Keywords: Bracteole; flower; flower-subtending bract; inflorescence; inhibitory field; pattern formation; prophyll; regulation of development; vasculature.
Figures
Similar articles
-
Diversity of flower structure and development correlates with inflorescence morphology in maples (Acer, Sapindaceae, Sapindales).Ann Bot. 2025 Jun 10:mcaf097. doi: 10.1093/aob/mcaf097. Online ahead of print. Ann Bot. 2025. PMID: 40492826
-
Towards an ontogenetic understanding of inflorescence diversity.Ann Bot. 2013 Nov;112(8):1523-42. doi: 10.1093/aob/mct009. Epub 2013 Feb 27. Ann Bot. 2013. PMID: 23445936 Free PMC article.
-
Flowers and inflorescences of the seagrass Posidonia (Posidoniaceae, Alismatales).Am J Bot. 2012 Oct;99(10):1592-608. doi: 10.3732/ajb.1200227. Epub 2012 Oct 2. Am J Bot. 2012. PMID: 23032814
-
Grass meristems II: inflorescence architecture, flower development and meristem fate.Plant Cell Physiol. 2013 Mar;54(3):313-24. doi: 10.1093/pcp/pct016. Epub 2013 Jan 31. Plant Cell Physiol. 2013. PMID: 23378448 Review.
-
Prophyll in Monocots: The Starting Point of Lateral Shoot Phyllotaxis.Front Plant Sci. 2022 Apr 13;13:855146. doi: 10.3389/fpls.2022.855146. eCollection 2022. Front Plant Sci. 2022. PMID: 35498710 Free PMC article.
Cited by
-
Two peculiar new species of Heteranthera Ruiz & Pavón (Pontederiaceae) from Brazil, with notes on inflorescence architecture in the family.PhytoKeys. 2017 Jun 28;(82):35-56. doi: 10.3897/phytokeys.82.13752. eCollection 2017. PhytoKeys. 2017. PMID: 28794681 Free PMC article.
-
Flower and Spikelet Construction in Rapateaceae (Poales).Front Plant Sci. 2022 Jan 27;12:813915. doi: 10.3389/fpls.2021.813915. eCollection 2021. Front Plant Sci. 2022. PMID: 35154210 Free PMC article.
-
Early inflorescence development in the grasses (Poaceae).Front Plant Sci. 2013 Jul 23;4:250. doi: 10.3389/fpls.2013.00250. eCollection 2013. Front Plant Sci. 2013. PMID: 23898335 Free PMC article.
-
Developmental Morphology and Anatomy Shed Light on Both Parallel and Convergent Evolution of the Umbellate Inflorescence in Monocots, Underlain by a New Variant of Metatopy.Front Plant Sci. 2022 Apr 29;13:873505. doi: 10.3389/fpls.2022.873505. eCollection 2022. Front Plant Sci. 2022. PMID: 35574142 Free PMC article.
-
COMPOSITUM 1 contributes to the architectural simplification of barley inflorescence via meristem identity signals.Nat Commun. 2020 Oct 12;11(1):5138. doi: 10.1038/s41467-020-18890-y. Nat Commun. 2020. PMID: 33046693 Free PMC article.
References
-
- Aloni R, Aloni E, Langhans M, Ullrich CI. Role of auxin in regulating Arabidopsis flower development. Planta. 2006;223:315–328. - PubMed
-
- Anderson CE. Some studies on the floral anatomy of the Liliales. Cornell University, Ithaca: NY; 1940. PhD Thesis.
-
- Arber A. Monocotyledons. A morphological study. Cambridge: Cambridge University Press; 1925.
-
- Azuma H, Tobe H. Molecular phylogenetic analyses of Tofieldiaceae (Alismatales): family circumscription and intergeneric relationships. Journal of Plant Research. 2010;124:349–357. - PubMed
-
- Barabé D, Bruneau A, Forest F, Lacroix C. The correlation between development of atypical bisexual flowers and phylogeny in the Aroideae (Araceae) Plant Systematics and Evolution. 2002;232:1–19.
