

Non-heritable genetics of human disease: spotlight on post-zygotic genetic variation acquired during lifetime
- PMID: 23172682
- PMCID: PMC3534255
- DOI: 10.1136/jmedgenet-2012-101322
Non-heritable genetics of human disease: spotlight on post-zygotic genetic variation acquired during lifetime
Abstract
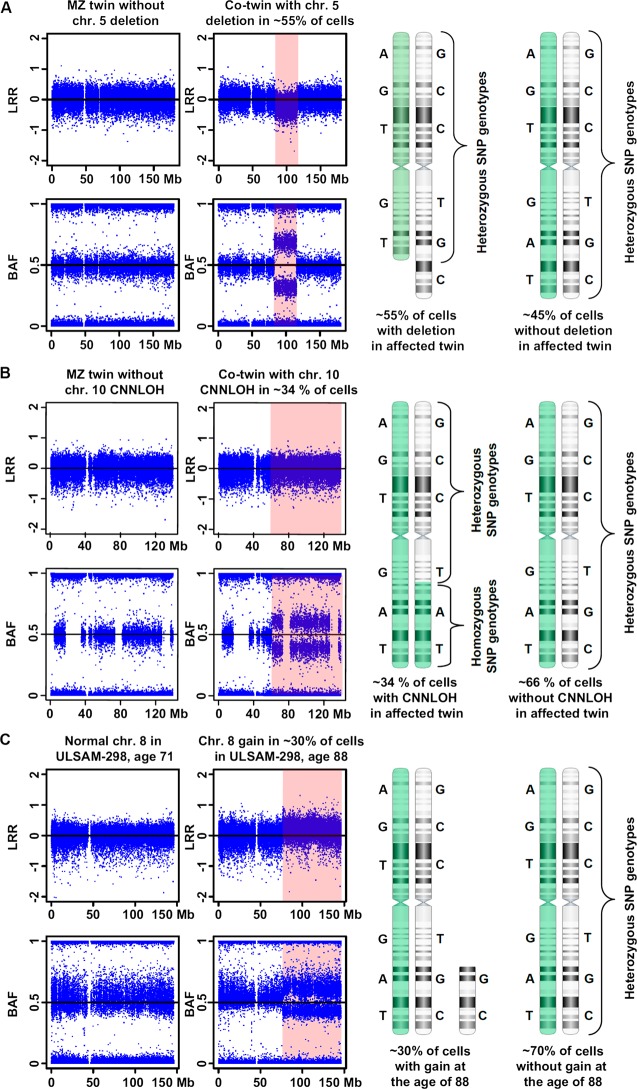
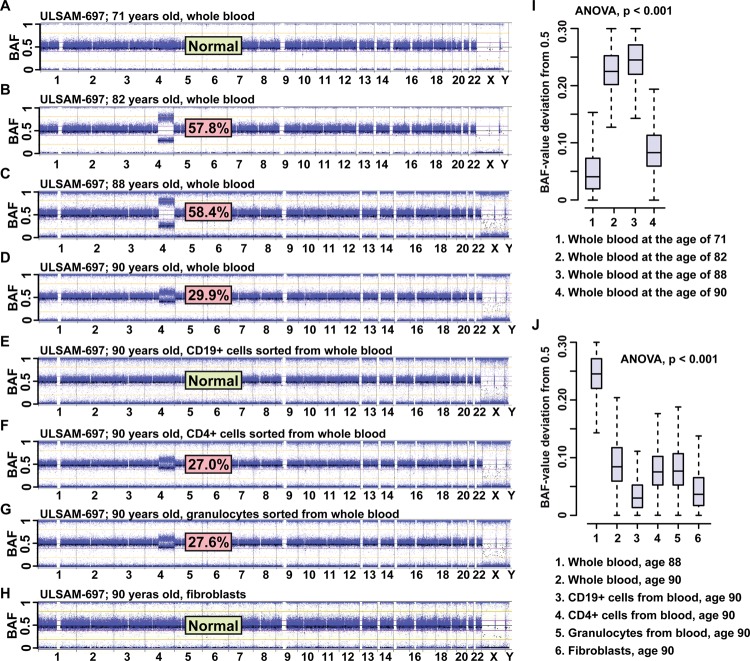
The heritability of most common, multifactorial diseases is rather modest and known genetic effects account for a small part of it. The remaining portion of disease aetiology has been conventionally ascribed to environmental effects, with an unknown part being stochastic. This review focuses on recent studies highlighting stochastic events of potentially great importance in human disease-the accumulation of post-zygotic structural aberrations with age in phenotypically normal humans. These findings are in agreement with a substantial mutational load predicted to occur during lifetime within the human soma. A major consequence of these results is that the genetic profile of a single tissue collected at one time point should be used with caution as a faithful portrait of other tissues from the same subject or the same tissue throughout life. Thus, the design of studies in human genetics interrogating a single sample per subject or applying lymphoblastoid cell lines may come into question. Sporadic disorders are common in medicine. We wish to stress the non-heritable genetic variation as a potentially important factor behind the development of sporadic diseases. Moreover, associations between post-zygotic mutations, clonal cell expansions and their relation to cancer predisposition are central in this context. Post-zygotic mutations are amenable to robust examination and are likely to explain a sizable part of non-heritable disease causality, which has routinely been thought of as synonymous with environmental factors. In view of the widespread accumulation of genetic aberrations with age and strong predictions of disease risk from such analyses, studies of post-zygotic mutations may be a fruitful approach for delineation of variants that are causative for common human disorders.
Figures
Republished in
-
Republished: Non-heritable genetics of human disease: spotlight on post-zygotic genetic variation acquired during lifetime.Postgrad Med J. 2013 Jul;89(1053):417-26. doi: 10.1136/postgradmedj-2012-101322rep. Postgrad Med J. 2013. PMID: 23781115 Free PMC article.
Similar articles
-
Republished: Non-heritable genetics of human disease: spotlight on post-zygotic genetic variation acquired during lifetime.Postgrad Med J. 2013 Jul;89(1053):417-26. doi: 10.1136/postgradmedj-2012-101322rep. Postgrad Med J. 2013. PMID: 23781115 Free PMC article.
-
Mosaicism in health and disease - clones picking up speed.Nat Rev Genet. 2017 Feb;18(2):128-142. doi: 10.1038/nrg.2016.145. Epub 2016 Dec 12. Nat Rev Genet. 2017. PMID: 27941868 Review.
-
Germline Mutations for Novel Candidate Predisposition Genes in Sporadic Schwannomatosis.Clin Orthop Relat Res. 2020 Nov;478(11):2442-2450. doi: 10.1097/CORR.0000000000001239. Clin Orthop Relat Res. 2020. PMID: 32281771 Free PMC article.
-
[Posttraumatic stress disorder (PTSD) as a consequence of the interaction between an individual genetic susceptibility, a traumatogenic event and a social context].Encephale. 2012 Oct;38(5):373-80. doi: 10.1016/j.encep.2011.12.003. Epub 2012 Jan 24. Encephale. 2012. PMID: 23062450 Review. French.
-
Detecting Familial Aggregation.Methods Mol Biol. 2017;1666:133-169. doi: 10.1007/978-1-4939-7274-6_8. Methods Mol Biol. 2017. PMID: 28980245
Cited by
-
Coherent somatic mutation in autoimmune disease.PLoS One. 2014 Jul 2;9(7):e101093. doi: 10.1371/journal.pone.0101093. eCollection 2014. PLoS One. 2014. PMID: 24988487 Free PMC article. Review.
-
Detection of SQSTM1/P392L post-zygotic mutations in Paget's disease of bone.Hum Genet. 2015 Jan;134(1):53-65. doi: 10.1007/s00439-014-1488-3. Epub 2014 Sep 21. Hum Genet. 2015. PMID: 25241215 Free PMC article.
-
A characterization of postzygotic mutations identified in monozygotic twins.Hum Mutat. 2018 Oct;39(10):1393-1401. doi: 10.1002/humu.23586. Epub 2018 Jul 18. Hum Mutat. 2018. PMID: 29980163 Free PMC article.
-
Mosaic Brain Aneuploidy in Mental Illnesses: An Association of Low-level Post-zygotic Aneuploidy with Schizophrenia and Comorbid Psychiatric Disorders.Curr Genomics. 2018 Apr;19(3):163-172. doi: 10.2174/1389202918666170717154340. Curr Genomics. 2018. PMID: 29606903 Free PMC article. Review.
-
Mutagenesis. Smoking is associated with mosaic loss of chromosome Y.Science. 2015 Jan 2;347(6217):81-3. doi: 10.1126/science.1262092. Epub 2014 Dec 4. Science. 2015. PMID: 25477213 Free PMC article.
References
-
- Forsberg L, Rasi C, Razzaghian H, Pakalapati G, Waite L, Thilbeault K Stanton, Ronowicz A, Wineinger N, Tiwari H, Boomsma D, Westerman M, Harris J, Lyle R, Essand M, Eriksson F, Strachan E, O'Hanlon T, Rider L, Miller F, Giedraitis V, Lannfelt L, Ingelsson M, Piotrowski A, Pedersen N, Absher D, Dumanski J. Age-related somatic structural changes in the nuclear genome of human blood cells. Am J Hum Genet 2012;90:217–28 - PMC - PubMed
-
- Laurie CC, Laurie CA, Rice K, Doheny KF, Zelnick LR, McHugh CP, Ling H, Hetrick KN, Pugh EW, Amos C, Wei Q, Wang LE, Lee JE, Barnes KC, Hansel NN, Mathias R, Daley D, Beaty TH, Scott AF, Ruczinski I, Scharpf RB, Bierut LJ, Hartz SM, Landi MT, Freedman ND, Goldin LR, Ginsburg D, Li J, Desch KC, Strom SS, Blot WJ, Signorello LB, Ingles SA, Chanock SJ, Berndt SI, Le Marchand L, Henderson BE, Monroe KR, Heit JA, de Andrade M, Armasu SM, Regnier C, Lowe WL, Hayes MG, Marazita ML, Feingold E, Murray JC, Melbye M, Feenstra B, Kang JH, Wiggs JL, Jarvik GP, McDavid AN, Seshan VE, Mirel DB, Crenshaw A, Sharopova N, Wise A, Shen J, Crosslin DR, Levine DM, Zheng X, Udren JI, Bennett S, Nelson SC, Gogarten SM, Conomos MP, Heagerty P, Manolio T, Pasquale LR, Haiman CA, Caporaso N, Weir BS. Detectable clonal mosaicism from birth to old age and its relationship to cancer. Nat Genet 2012;44:642–50 - PMC - PubMed
-
- Jacobs KB, Yeager M, Zhou W, Wacholder S, Wang Z, Rodriguez-Santiago B, Hutchinson A, Deng X, Liu C, Horner MJ, Cullen M, Epstein CG, Burdett L, Dean MC, Chatterjee N, Sampson J, Chung CC, Kovaks J, Gapstur SM, Stevens VL, Teras LT, Gaudet MM, Albanes D, Weinstein SJ, Virtamo J, Taylor PR, Freedman ND, Abnet CC, Goldstein AM, Hu N, Yu K, Yuan JM, Liao L, Ding T, Qiao YL, Gao YT, Koh WP, Xiang YB, Tang ZZ, Fan JH, Aldrich MC, Amos C, Blot WJ, Bock CH, Gillanders EM, Harris CC, Haiman CA, Henderson BE, Kolonel LN, Le Marchand L, McNeill LH, Rybicki BA, Schwartz AG, Signorello LB, Spitz MR, Wiencke JK, Wrensch M, Wu X, Zanetti KA, Ziegler RG, Figueroa JD, Garcia-Closas M, Malats N, Marenne G, Prokunina-Olsson L, Baris D, Schwenn M, Johnson A, Landi MT, Goldin L, Consonni D, Bertazzi PA, Rotunno M, Rajaraman P, Andersson U, Freeman LE Beane, Berg CD, Buring JE, Butler MA, Carreon T, Feychting M, Ahlbom A, Gaziano JM, Giles GG, Hallmans G, Hankinson SE, Hartge P, Henriksson R, Inskip PD, Johansen C, Landgren A, McKean-Cowdin R, Michaud DS, Melin BS, Peters U, Ruder AM, Sesso HD, Severi G, Shu XO, Visvanathan K, White E, Wolk A, Zeleniuch-Jacquotte A, Zheng W, Silverman DT, Kogevinas M, Gonzalez JR, Villa O, Li D, Duell EJ, Risch HA, Olson SH, Kooperberg C, Wolpin BM, Jiao L, Hassan M, Wheeler W, Arslan AA, Bueno-de-Mesquita HB, Fuchs CS, Gallinger S, Gross MD, Holly EA, Klein AP, LaCroix A, Mandelson MT, Petersen G, Boutron-Ruault MC, Bracci PM, Canzian F, Chang K, Cotterchio M, Giovannucci EL, Goggins M, Bolton JA Hoffman, Jenab M, Khaw KT, Krogh V, Kurtz RC, McWilliams RR, Mendelsohn JB, Rabe KG, Riboli E, Tjonneland A, Tobias GS, Trichopoulos D, Elena JW, Yu H, Amundadottir L, Stolzenberg-Solomon RZ, Kraft P, Schumacher F, Stram D, Savage SA, Mirabello L, Andrulis IL, Wunder JS, Garcia A Patino, Sierrasesumaga L, Barkauskas DA, Gorlick RG, Purdue M, Chow WH, Moore LE, Schwartz KL, Davis FG, Hsing AW, Berndt SI, Black A, Wentzensen N, Brinton LA, Lissowska J, Peplonska B, McGlynn KA, Cook MB, Graubard BI, Kratz CP, Greene MH, Erickson RL, Hunter DJ, Thomas G, Hoover RN, Real FX, Fraumeni JF, Jr, Caporaso NE, Tucker M, Rothman N, Perez-Jurado LA, Chanock SJ. Detectable clonal mosaicism and its relationship to aging and cancer. Nat Genet 2012;44:651–8 - PMC - PubMed
-
- Maher B. Personal genomes: The case of the missing heritability. Nature 2008;456:18–21 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources