

Evidence for autoregulation and cell signaling pathway regulation from genome-wide binding of the Drosophila retinoblastoma protein
- PMID: 23173097
- PMCID: PMC3484676
- DOI: 10.1534/g3.112.004424
Evidence for autoregulation and cell signaling pathway regulation from genome-wide binding of the Drosophila retinoblastoma protein
Abstract
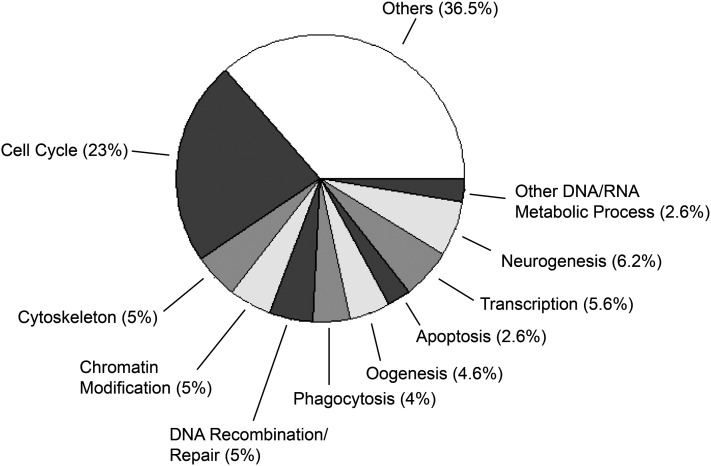
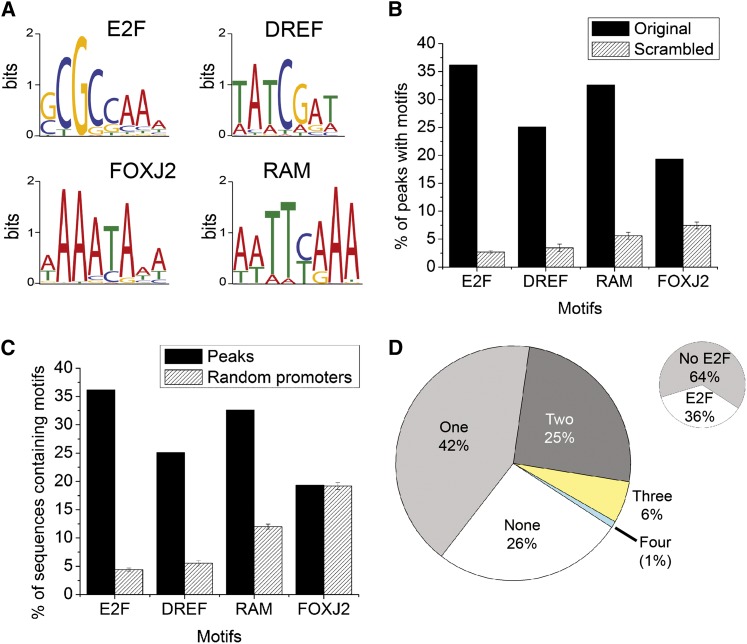
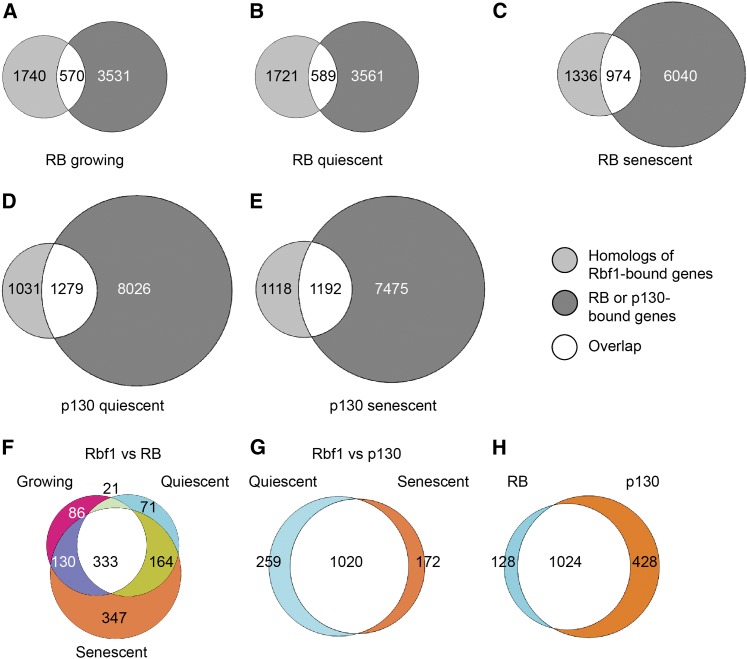
The retinoblastoma (RB) tumor suppressor protein is a transcriptional cofactor with essential roles in cell cycle and development. Physical and functional targets of RB and its paralogs p107/p130 have been studied largely in cultured cells, but the full biological context of this family of proteins' activities will likely be revealed only in whole organismal studies. To identify direct targets of the major Drosophila RB counterpart in a developmental context, we carried out ChIP-Seq analysis of Rbf1 in the embryo. The association of the protein with promoters is developmentally controlled; early promoter access is globally inhibited, whereas later in development Rbf1 is found to associate with promoter-proximal regions of approximately 2000 genes. In addition to conserved cell-cycle-related genes, a wholly unexpected finding was that Rbf1 targets many components of the insulin, Hippo, JAK/STAT, Notch, and other conserved signaling pathways. Rbf1 may thus directly affect output of these essential growth-control and differentiation pathways by regulation of expression of receptors, kinases and downstream effectors. Rbf1 was also found to target multiple levels of its own regulatory hierarchy. Bioinformatic analysis indicates that different classes of genes exhibit distinct constellations of motifs associated with the Rbf1-bound regions, suggesting that the context of Rbf1 recruitment may vary within the Rbf1 regulon. Many of these targeted genes are bound by Rbf1 homologs in human cells, indicating that a conserved role of RB proteins may be to adjust the set point of interlinked signaling networks essential for growth and development.
Keywords: Drosophila; Rbf1; cell-cycle; retinoblastoma.
Figures
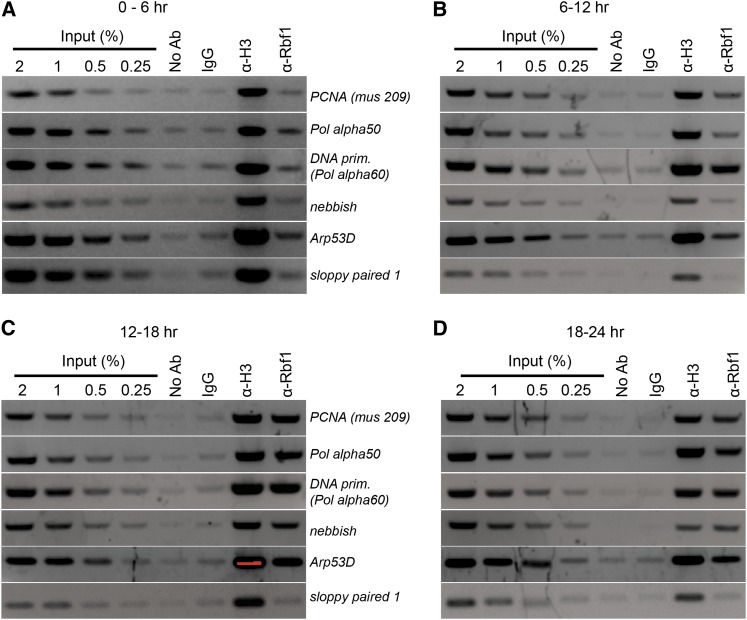
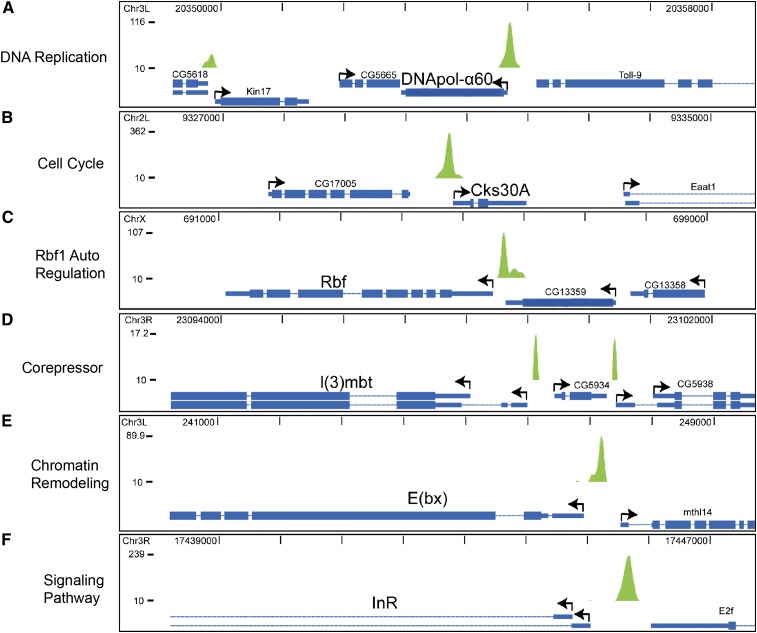
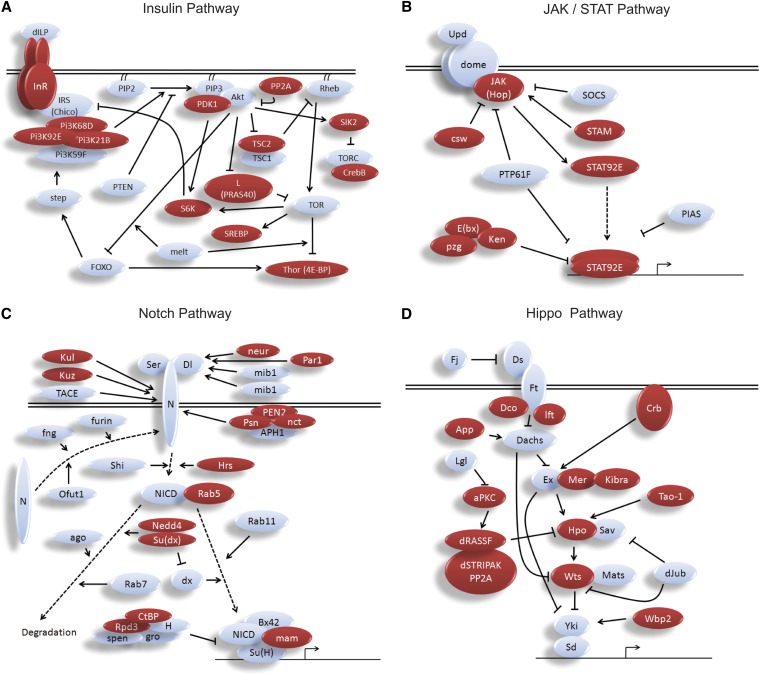
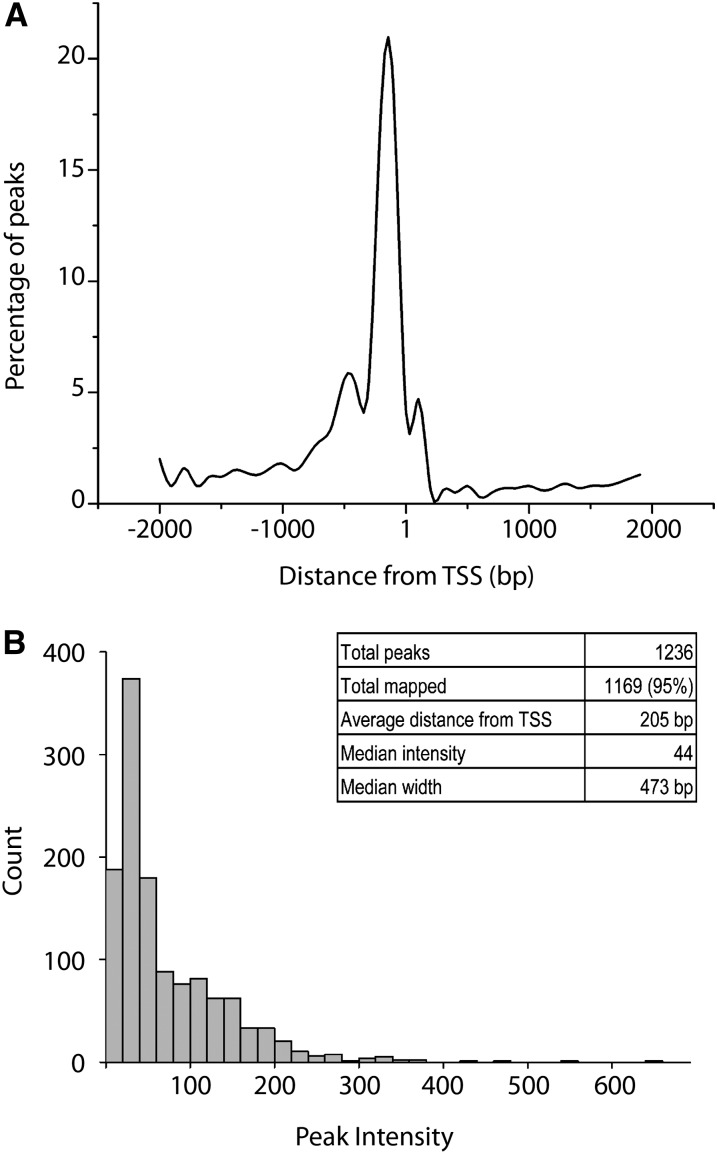

References
-
- Andersen E. C., Lu X., Horvitz H. R., 2006. C. elegans ISWI and NURF301 antagonize an Rb-like pathway in the determination of multiple cell fates. Development 133: 2695–2704 - PubMed
-
- Bailey T. L., Elkan C., 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 2: 28–36 - PubMed
-
- Batsche E., Moschopoulos P., Desroches J., Bilodeau S., Drouin J., 2005. Retinoblastoma and the related pocket protein p107 act as coactivators of NeuroD1 to enhance gene transcription. J. Biol. Chem. 280: 16088–16095 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases