

Functional relationships for investigating cognitive processes
- PMID: 23174335
- PMCID: PMC3582779
- DOI: 10.1016/j.beproc.2012.11.003
Functional relationships for investigating cognitive processes
Abstract
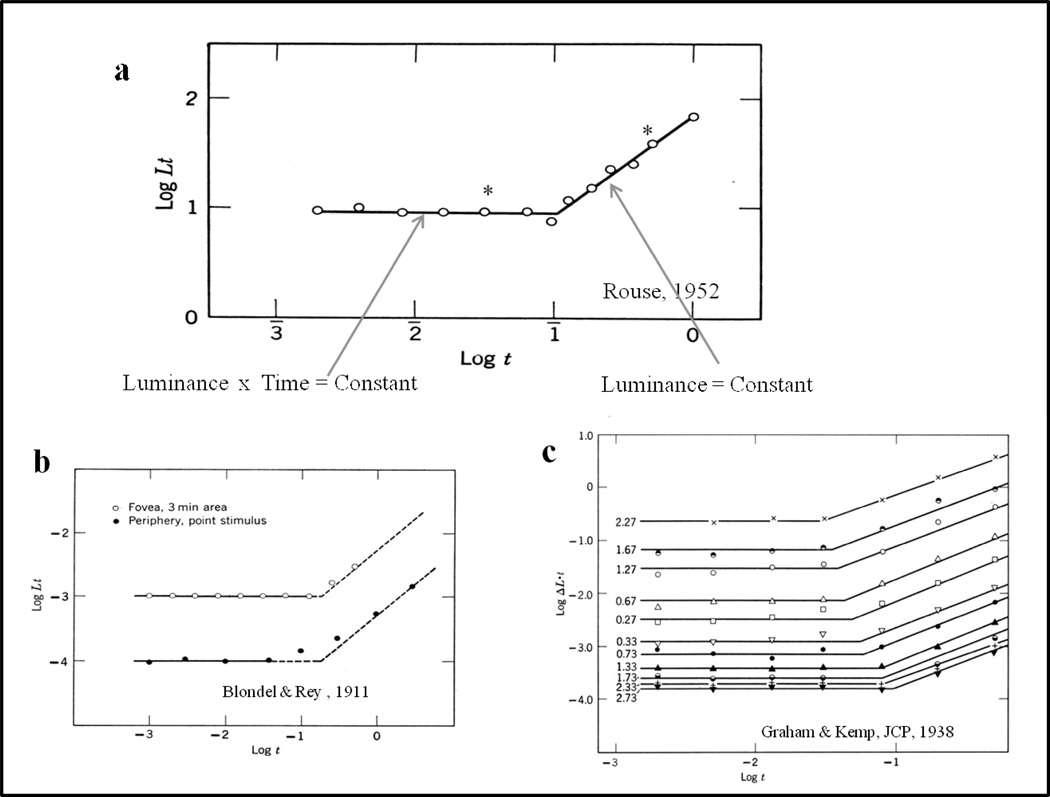
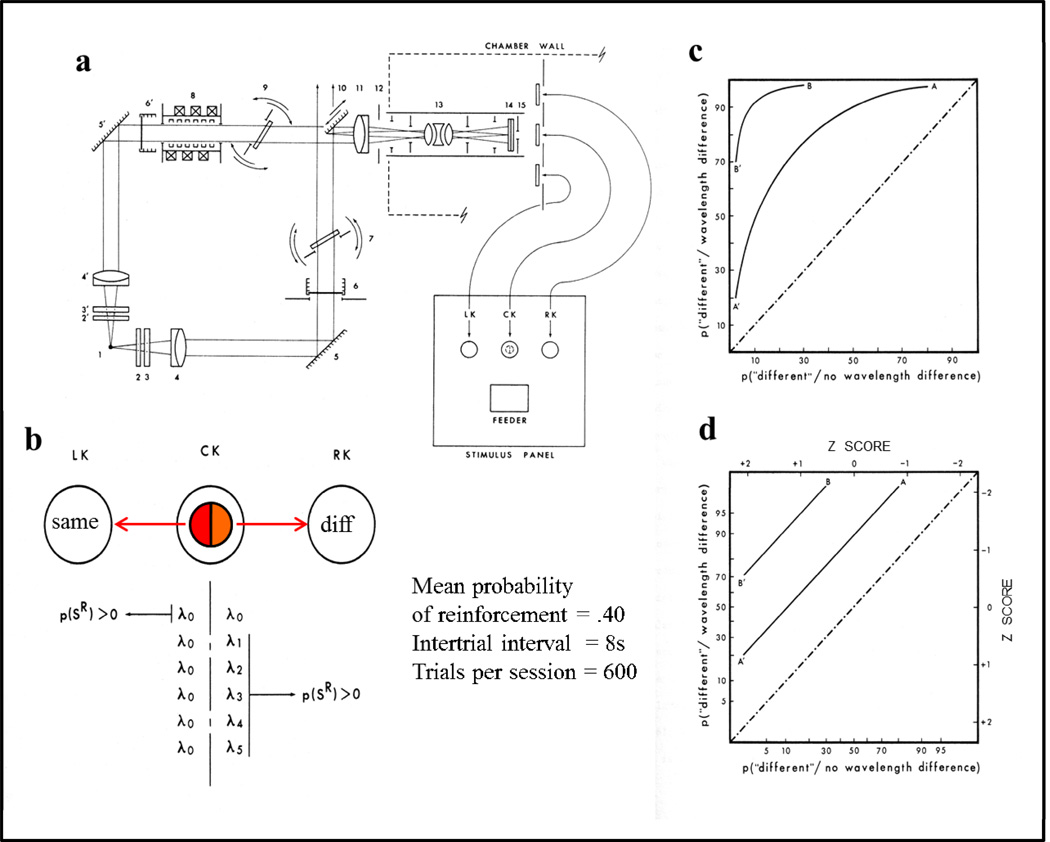
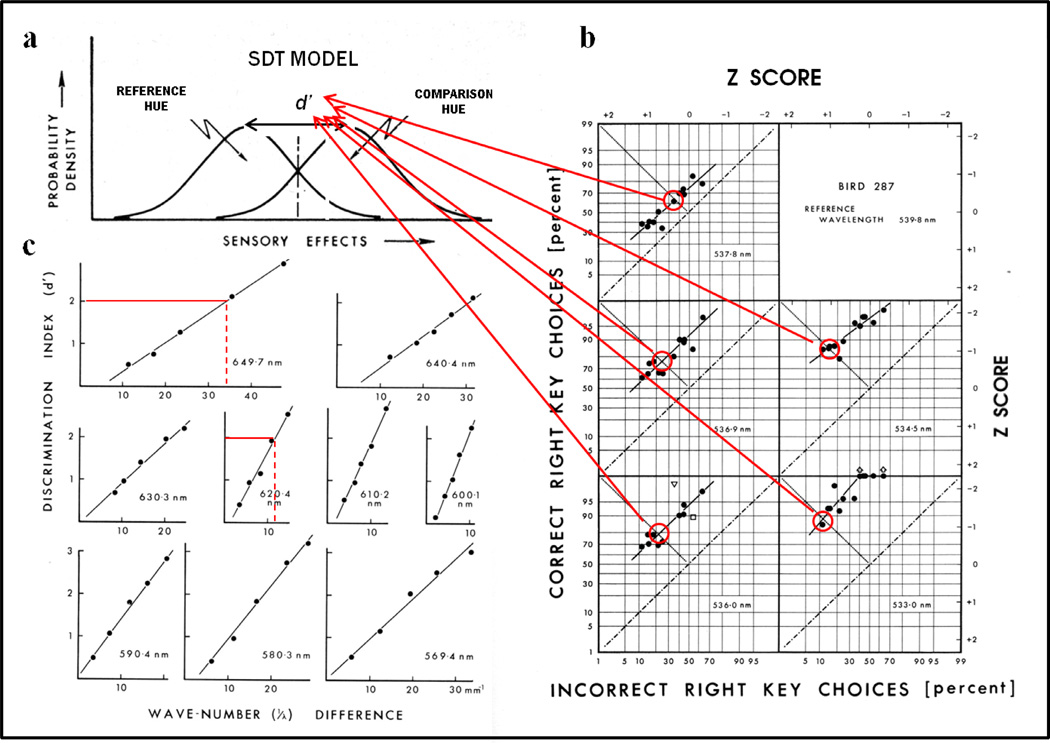
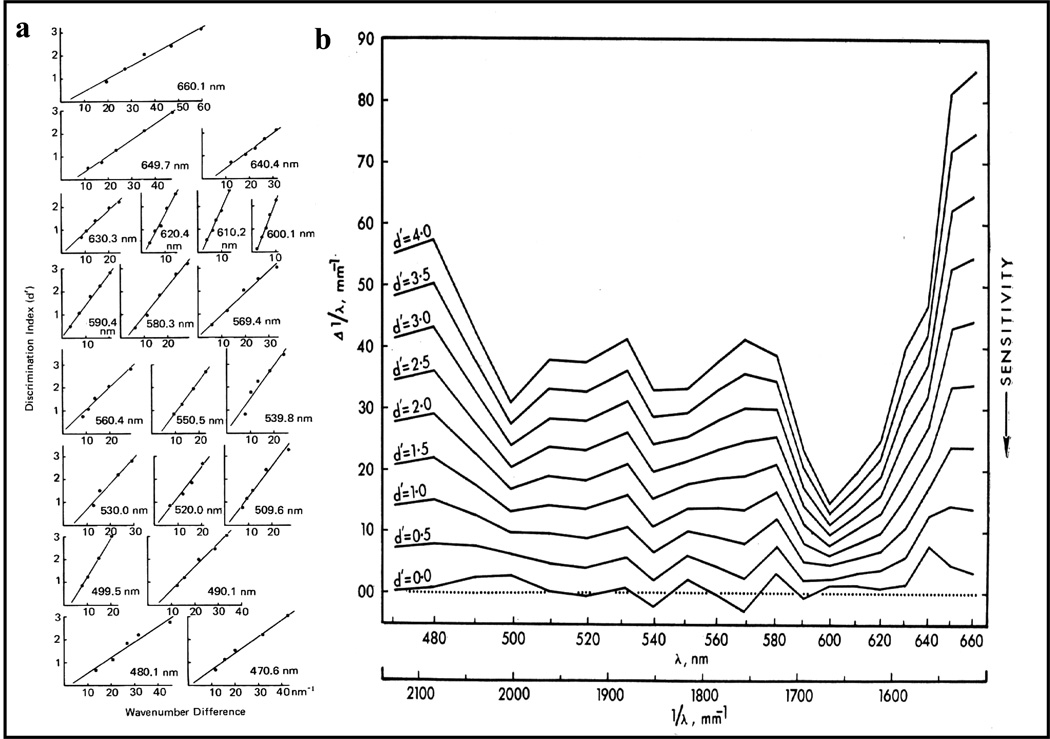
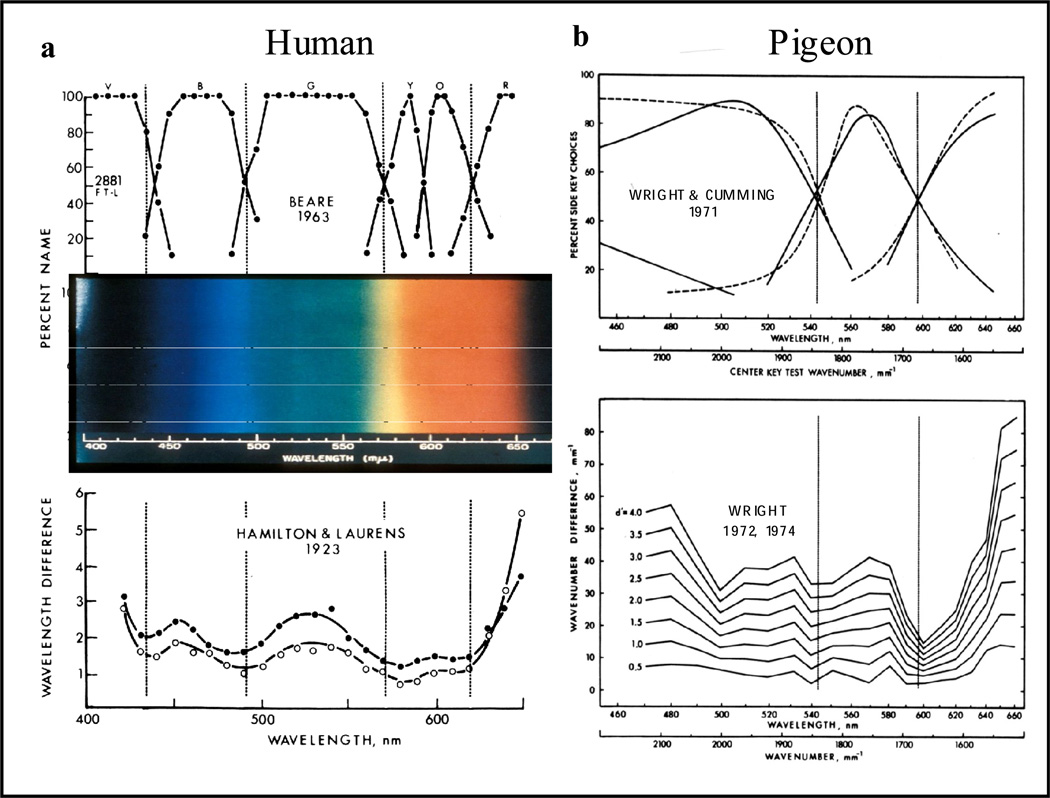
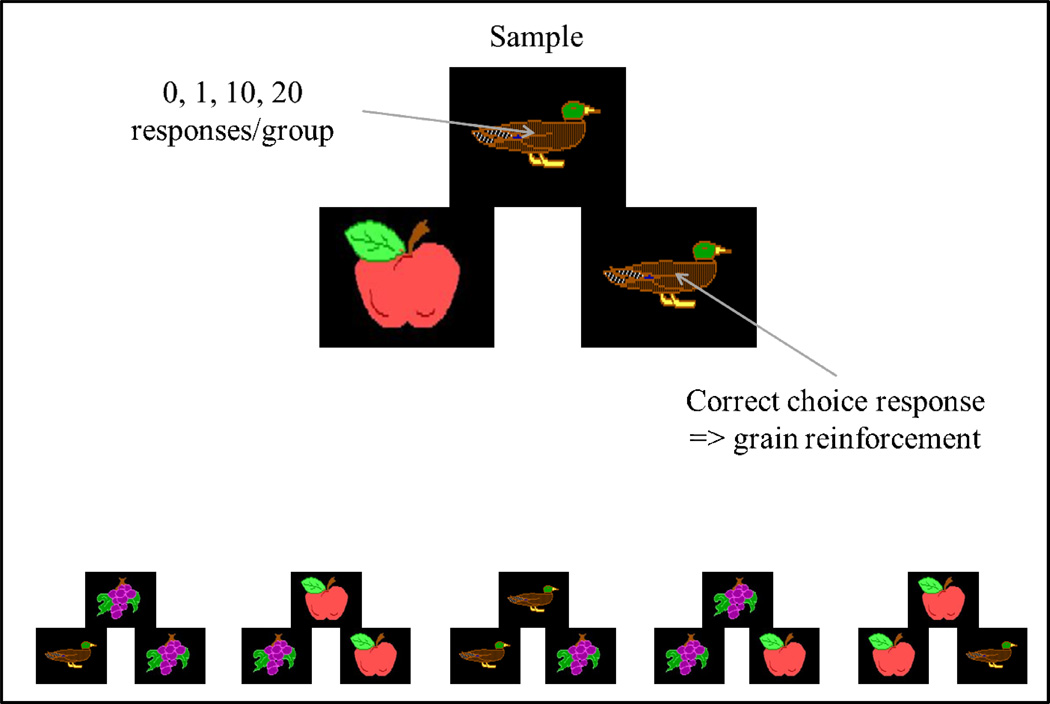
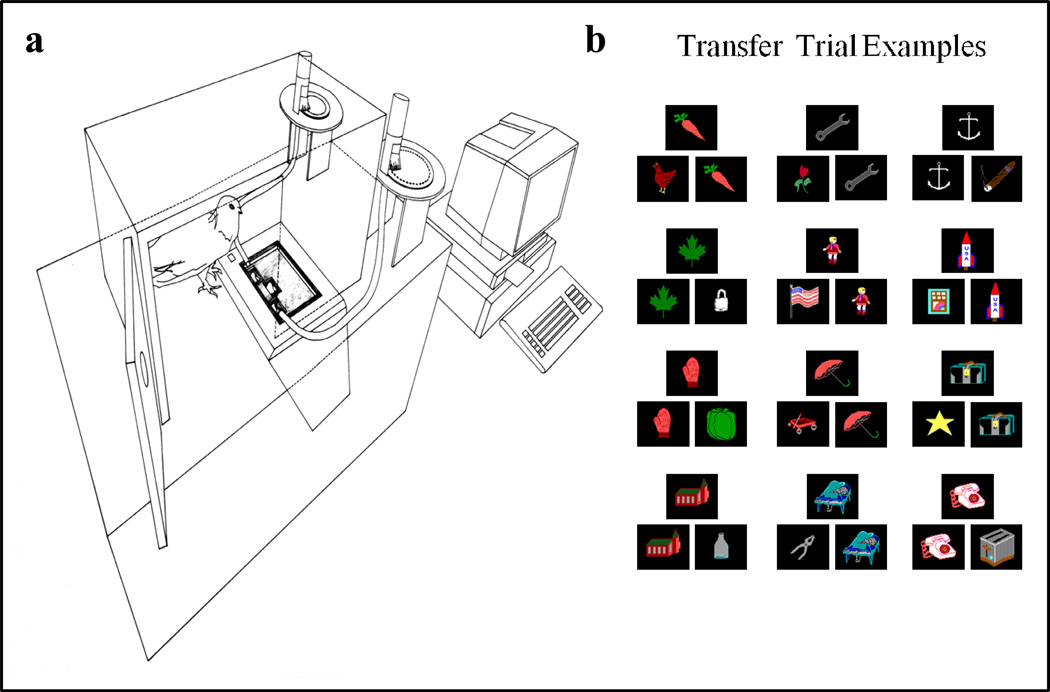
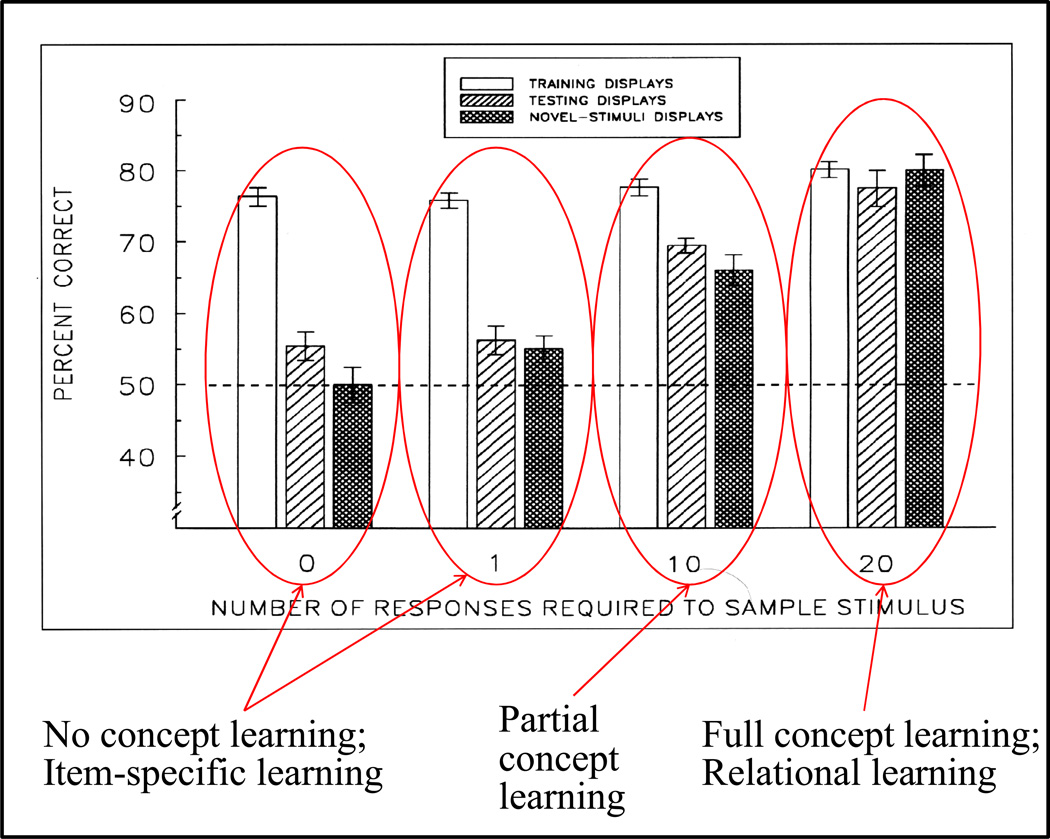
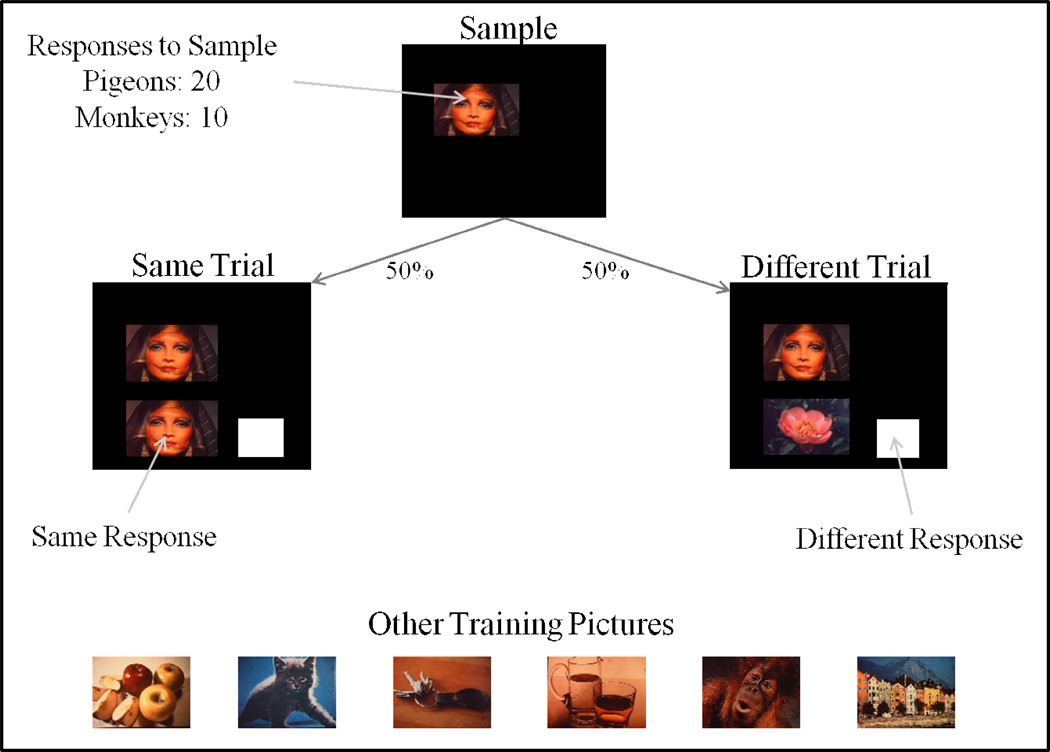
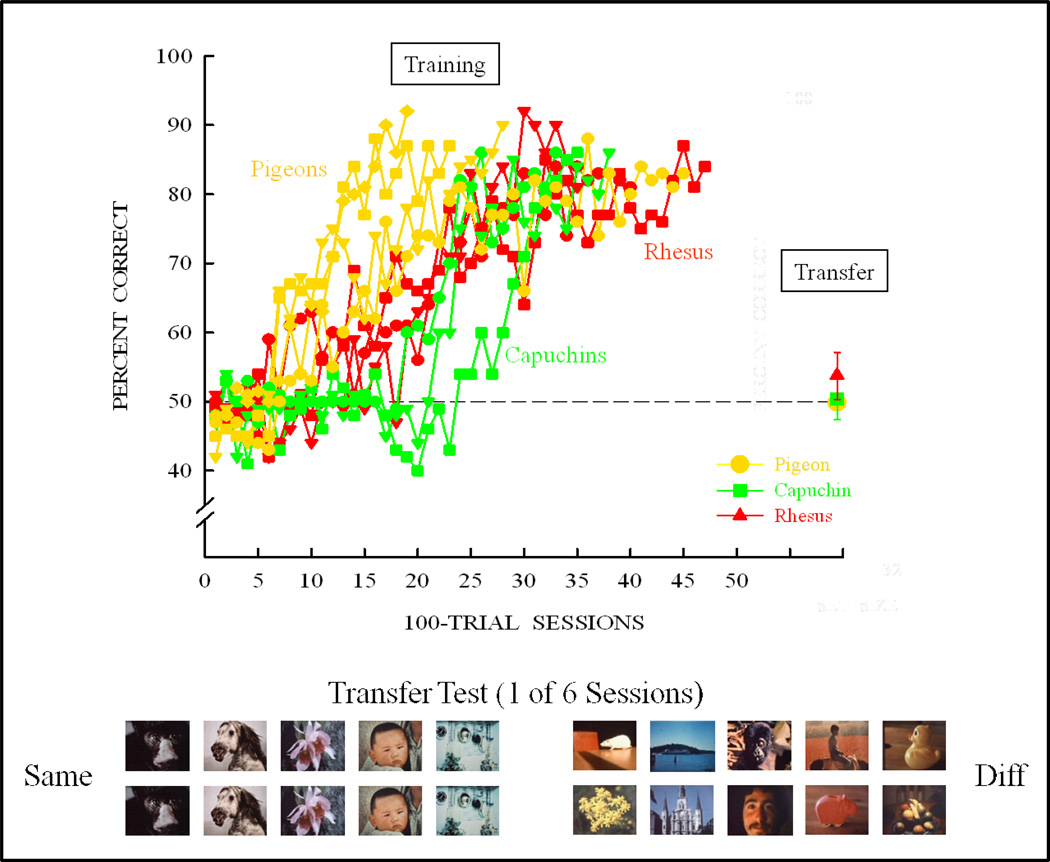
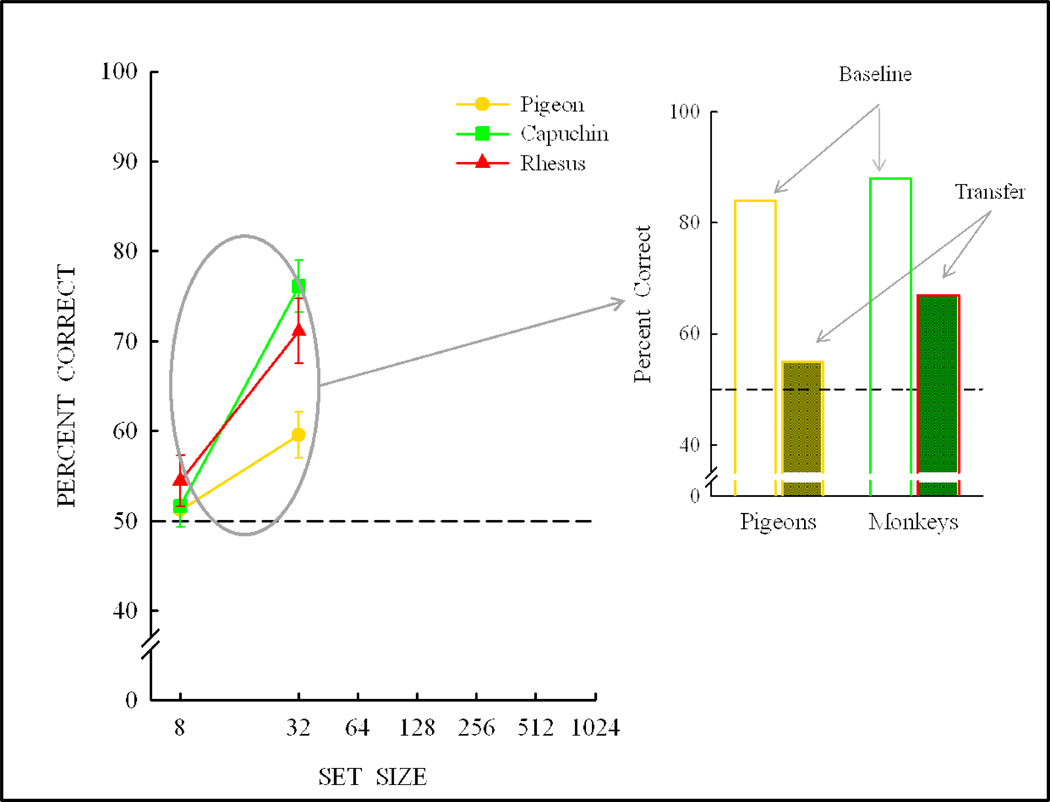
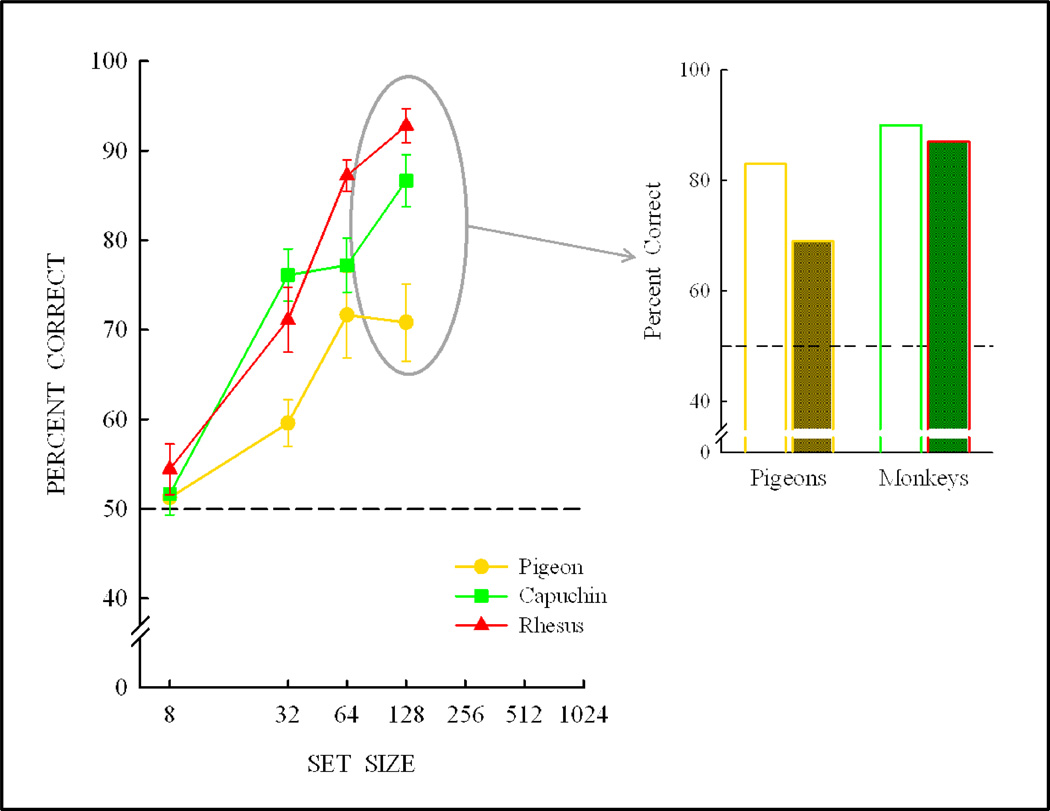
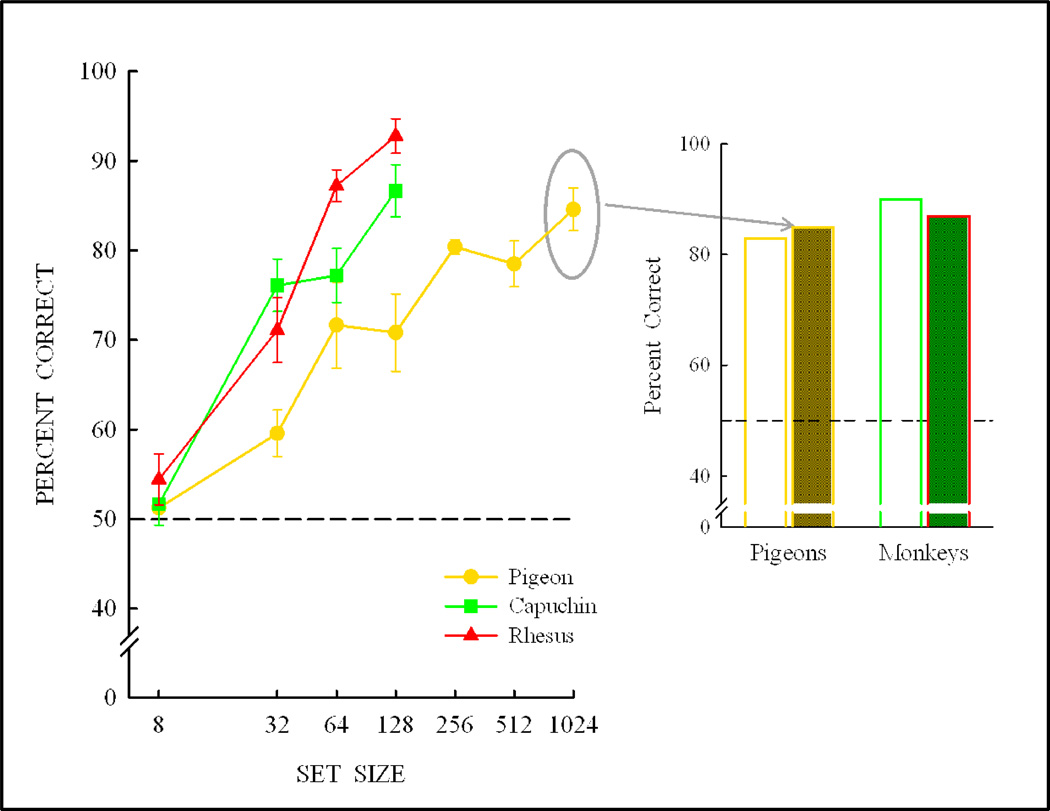
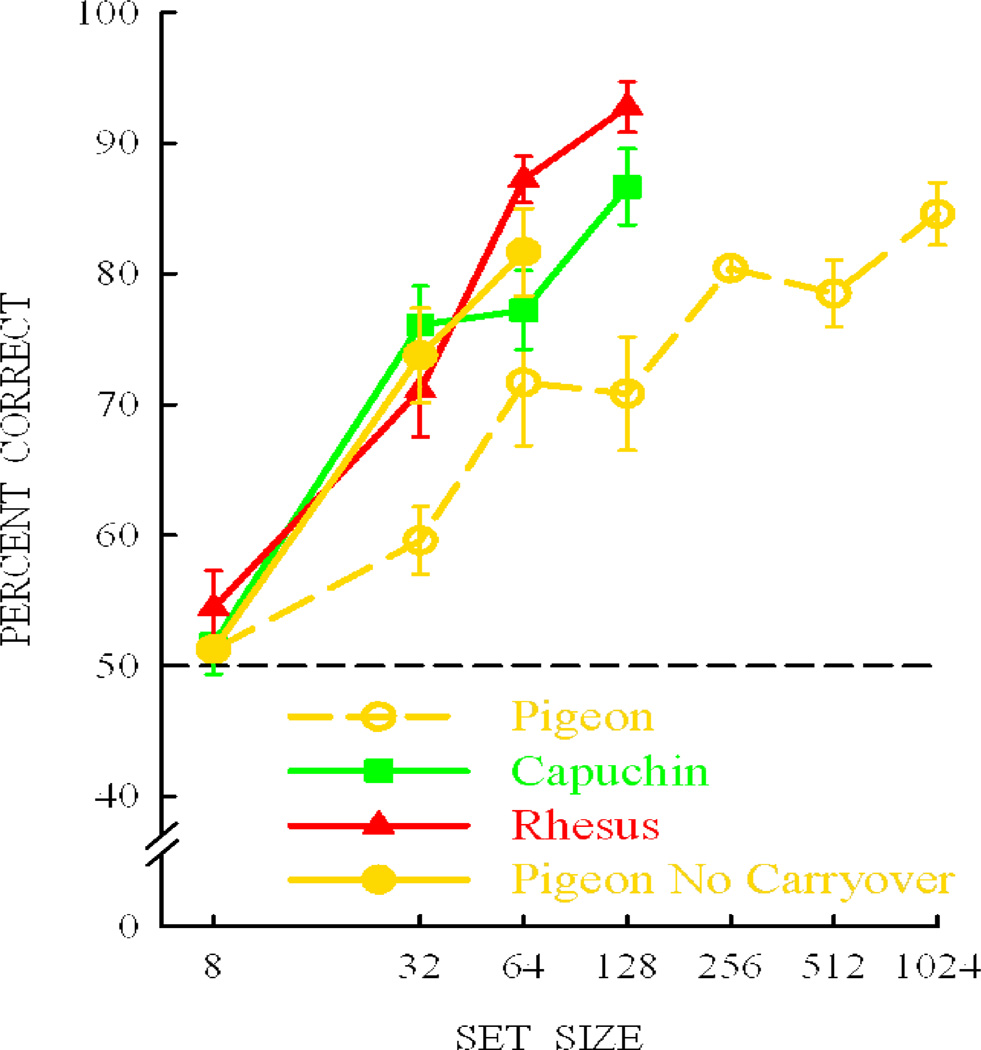
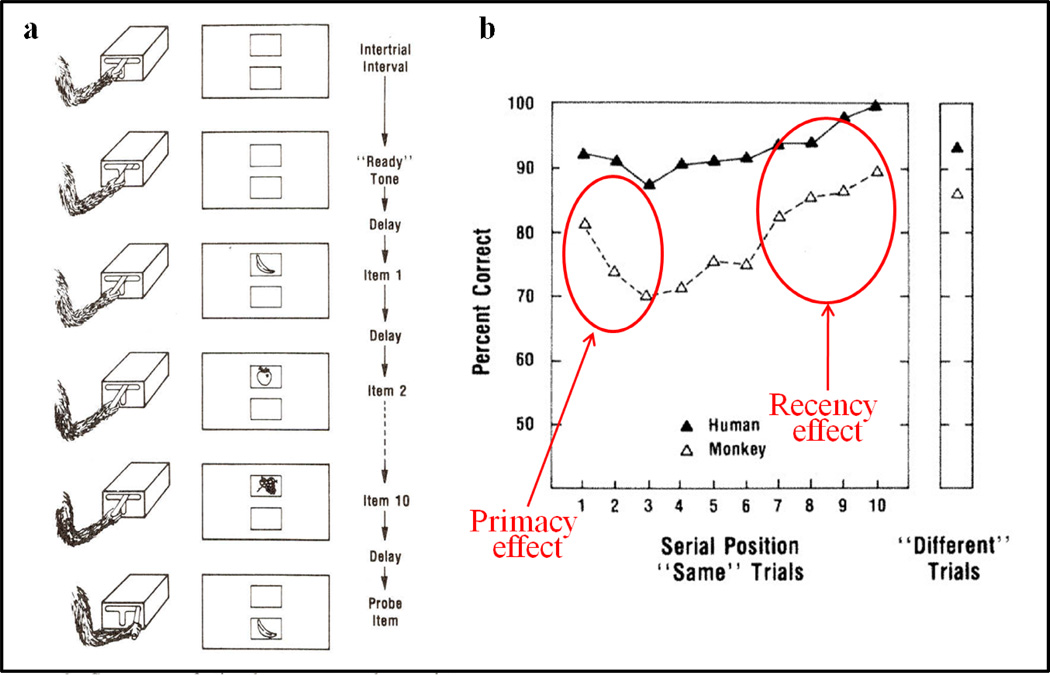
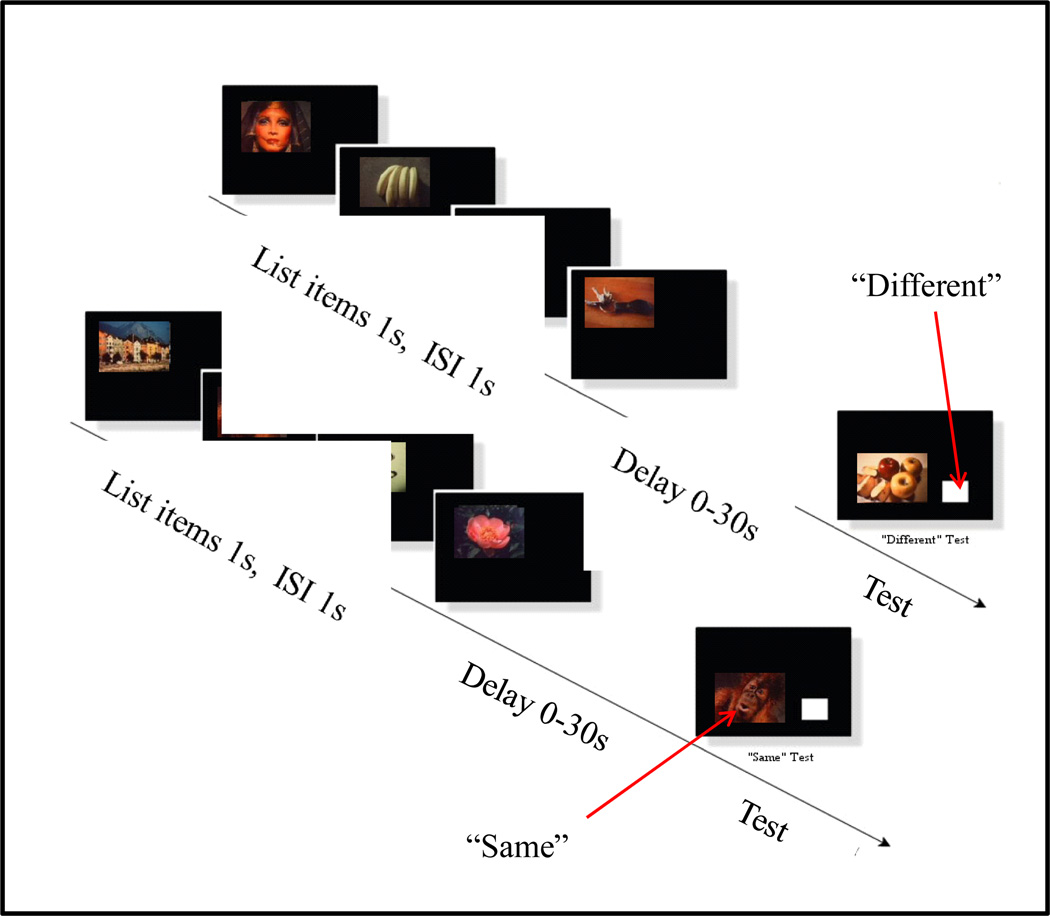
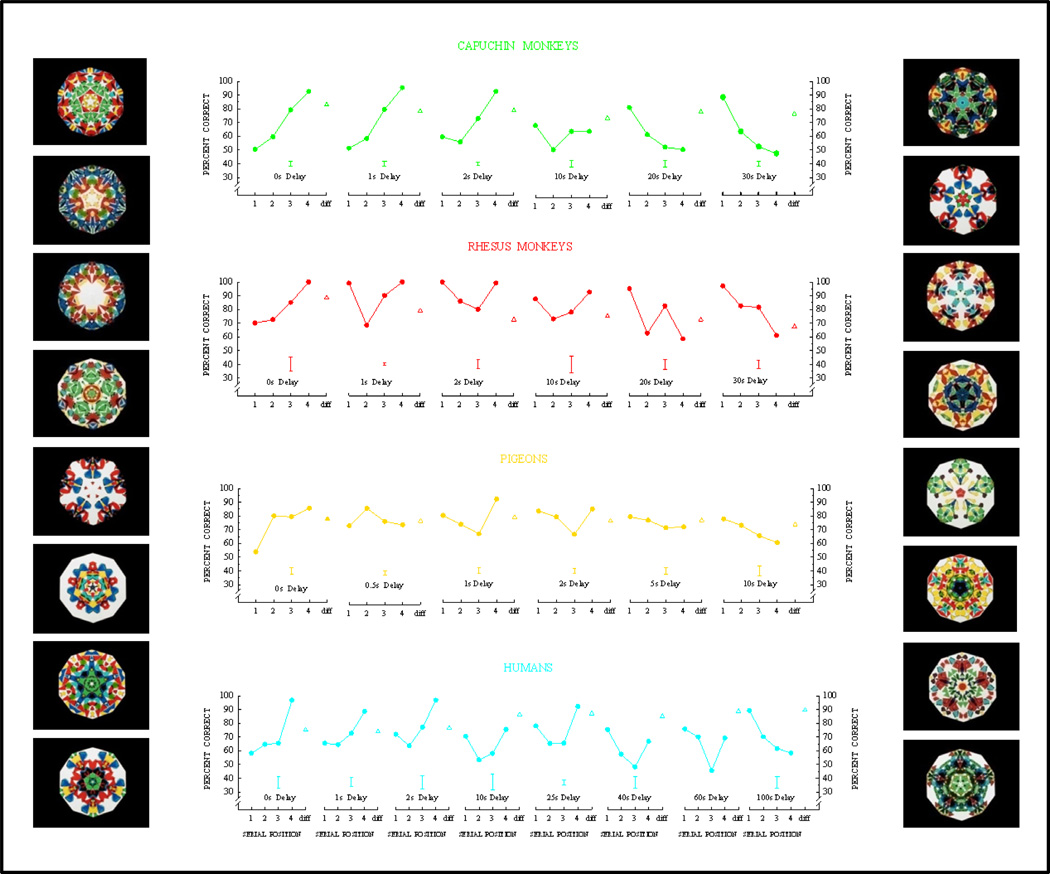
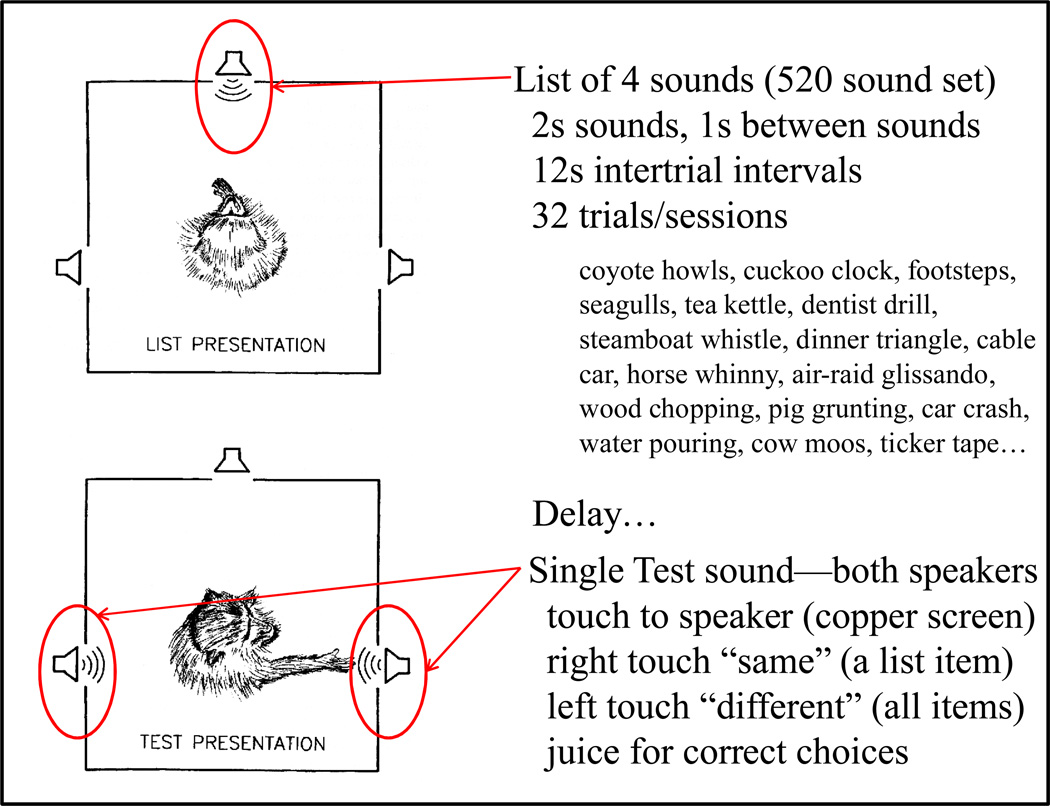
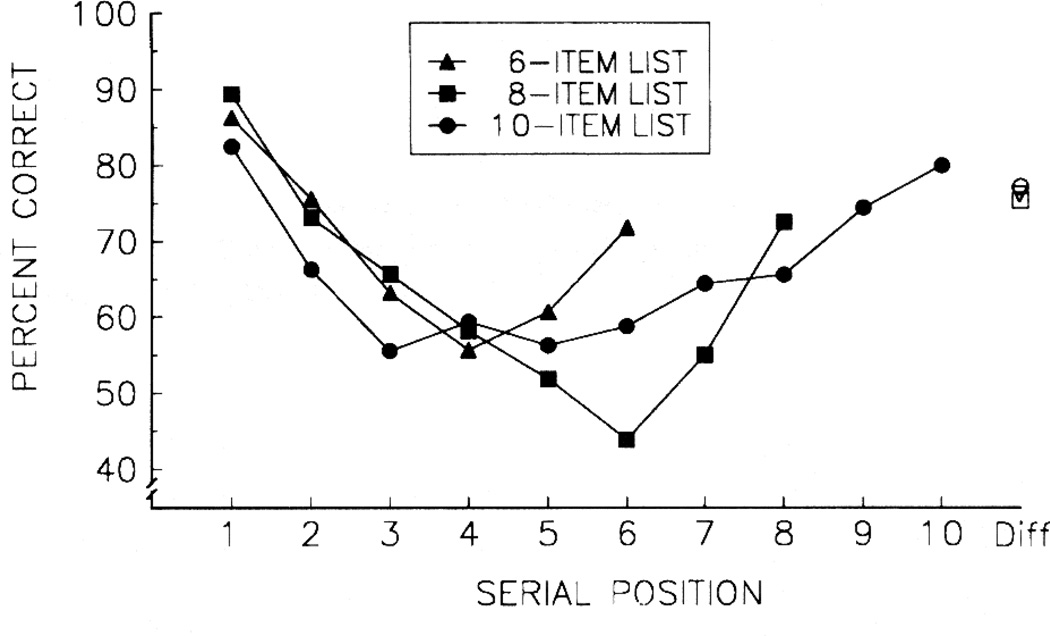
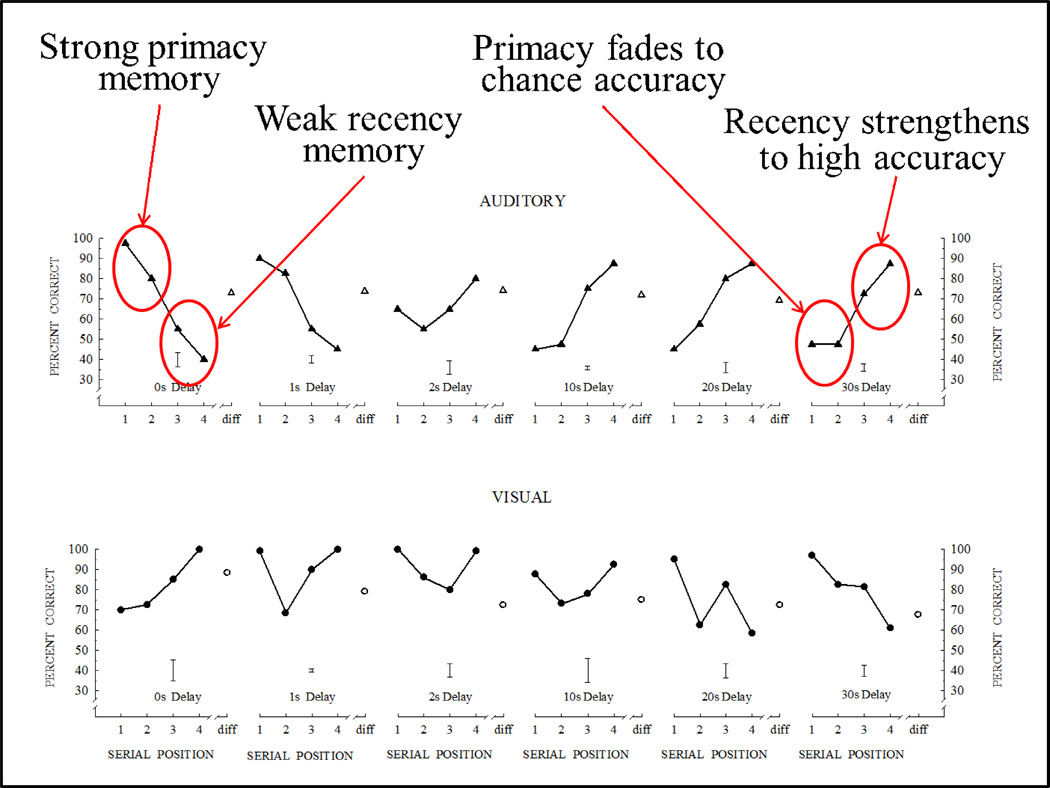
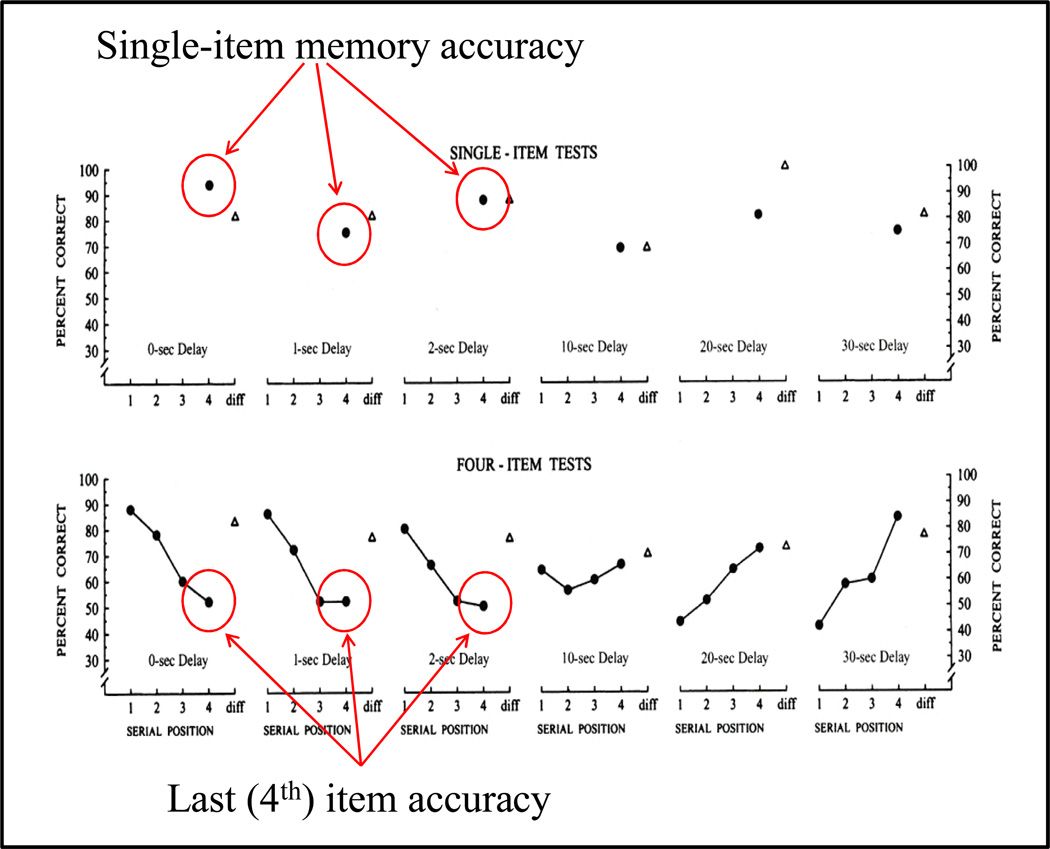
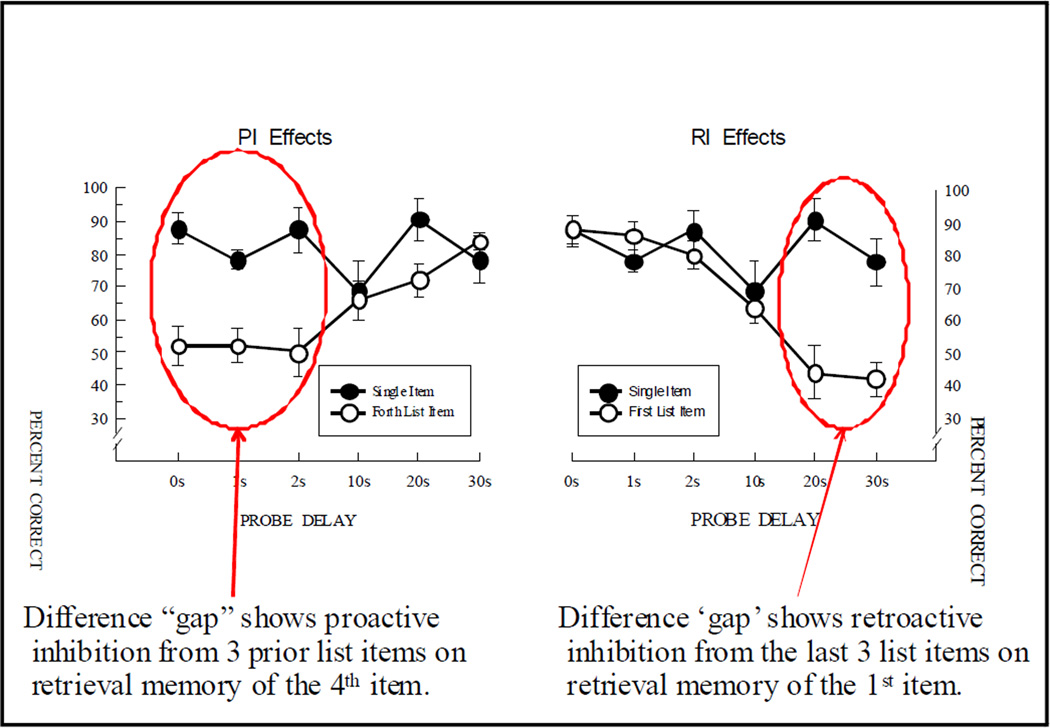
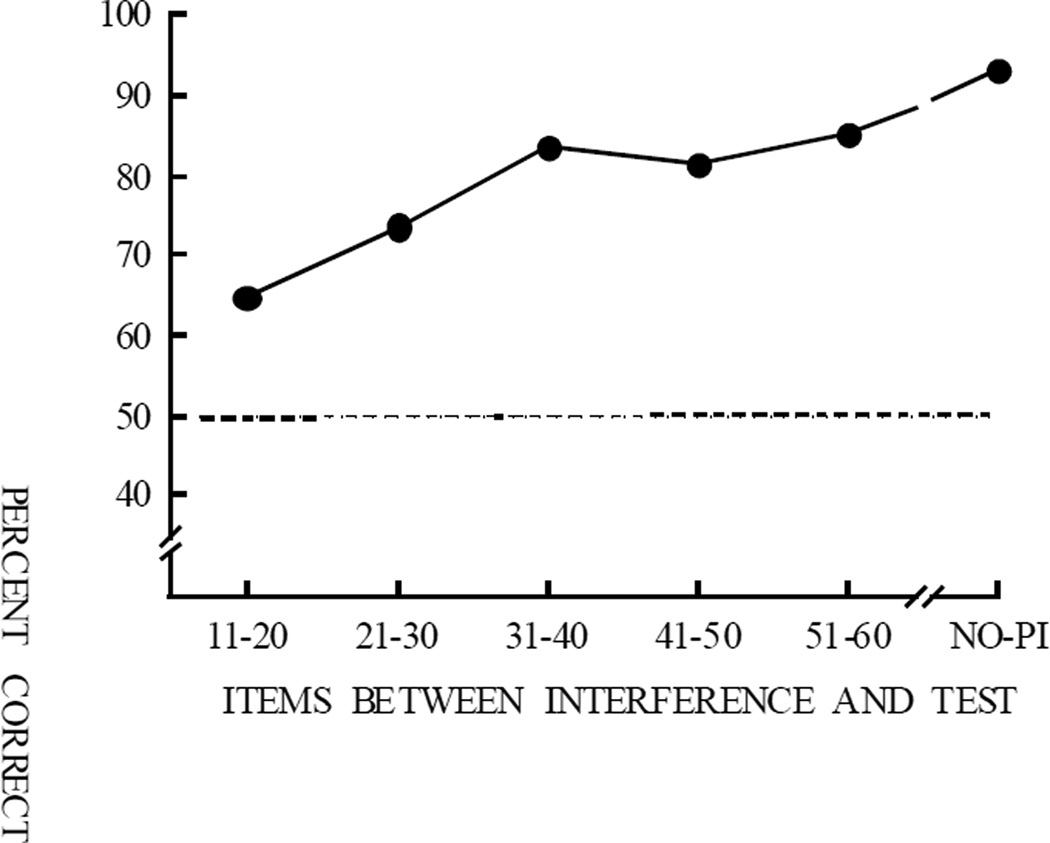
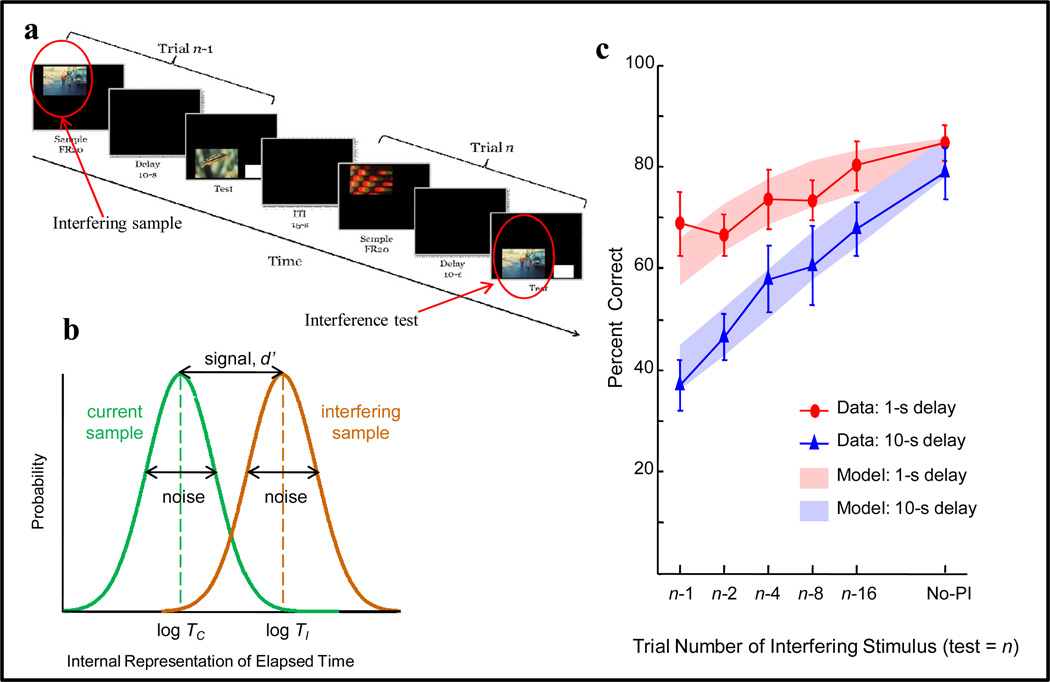
Functional relationships (from systematic manipulation of critical variables) are advocated for revealing fundamental processes of (comparative) cognition-through examples from my work in psychophysics, learning, and memory. Functional relationships for pigeon wavelength (hue) discrimination revealed best discrimination at the spectral points of hue transition for pigeons-a correspondence (i.e., functional relationship) similar to that for humans. Functional relationships for learning revealed: Item-specific or relational learning in matching to sample as a function of the pigeons' sample-response requirement, and same/different abstract-concept learning as a function of the training set size for rhesus monkeys, capuchin monkeys, and pigeons. Functional relationships for visual memory revealed serial position functions (a 1st order functional relationship) that changed systematically with retention delay (a 2nd order relationship) for pigeons, capuchin monkeys, rhesus monkeys, and humans. Functional relationships for rhesus-monkey auditory memory also revealed systematic changes in serial position functions with delay, but these changes were opposite to those for visual memory. Functional relationships for proactive interference revealed interference that varied as a function of a ratio of delay times. Functional relationships for change detection memory revealed (qualitative) similarities and (quantitative) differences in human and monkey visual short-term memory as a function of the number of memory items. It is concluded that these findings were made possible by varying critical variables over a substantial portion of the manipulable range to generate functions and derive relationships.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures




Comment in
-
Functional relationships.Behav Processes. 2013 Feb;93:1-3. doi: 10.1016/j.beproc.2013.01.004. Epub 2013 Jan 24. Behav Processes. 2013. PMID: 23353725 No abstract available.
References
-
- Alvarez GA, Cavanagh P. The Capacity of Visual Short-Term Memory is Set Both by Visual Information Load and by Number of Objects. Psychological Science. 2004;15:106–111. - PubMed
-
- Atkinson RC, Shiffrin RM. Human memory: A proposed system and its control processes. In: Spence KW, Spence JT, editors. The psychology of learning and motivation. Vol. 2. New York: Academic Press; 1968. pp. 89–105.
-
- Bower GH. An association model for response and training variables in paired-associate learning. Psychological Review. 1962;69:34–53. - PubMed
-
- Bower GH. Neal E. Miller. Proceedings of the American Philosophical Society. 2011;155:358–365.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
