

Mis-expression of the BK K(+) channel disrupts suprachiasmatic nucleus circuit rhythmicity and alters clock-controlled behavior
- PMID: 23174562
- PMCID: PMC3566534
- DOI: 10.1152/ajpcell.00302.2012
Mis-expression of the BK K(+) channel disrupts suprachiasmatic nucleus circuit rhythmicity and alters clock-controlled behavior
Abstract
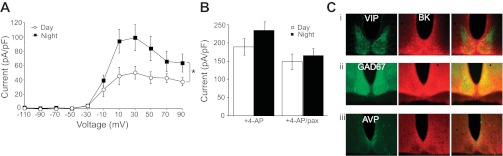
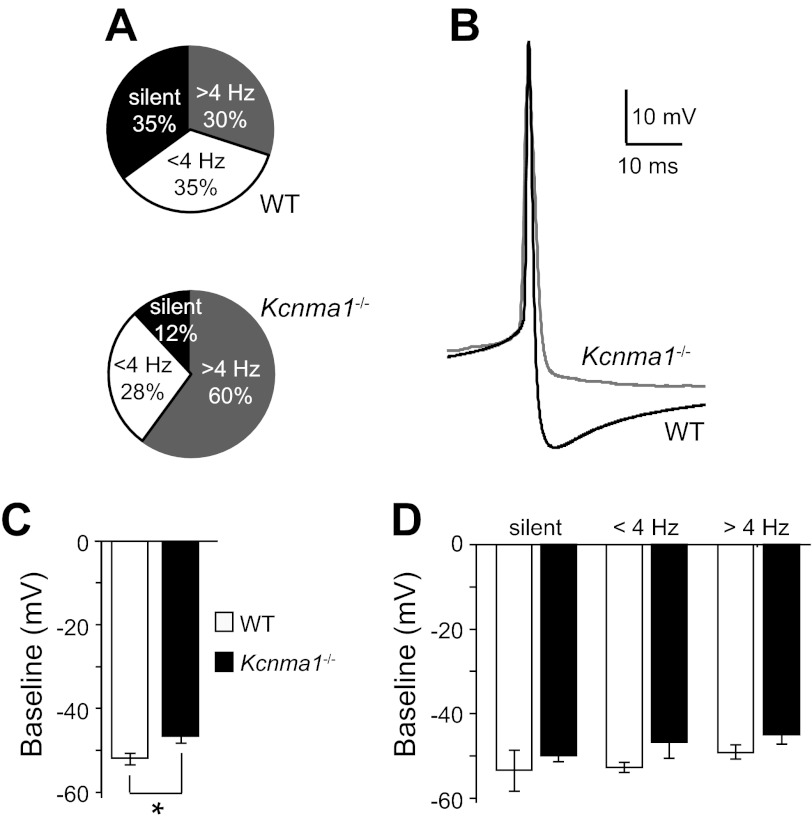
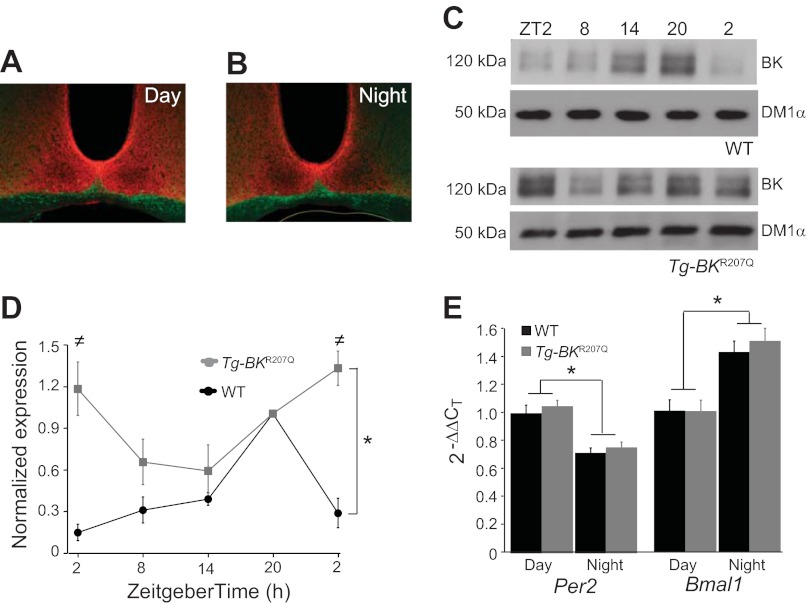
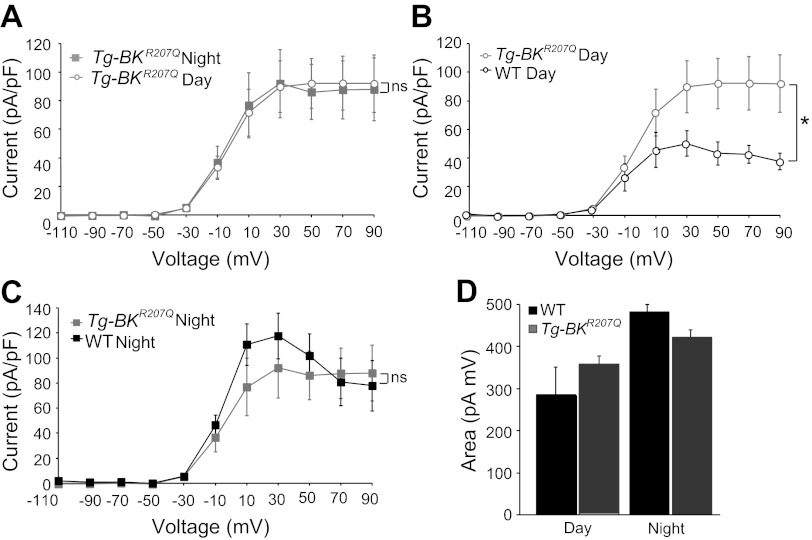
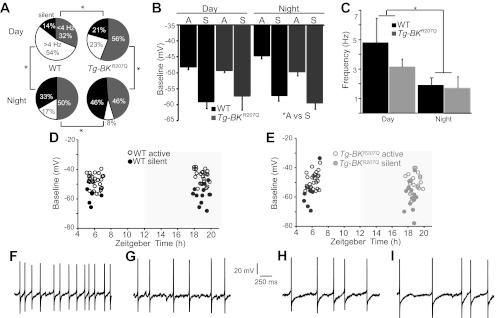
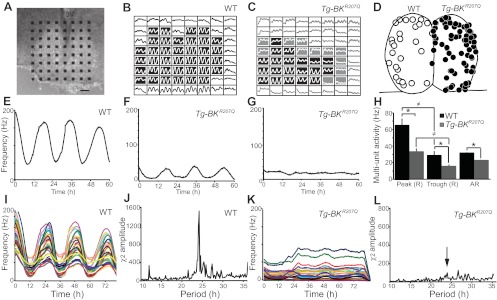
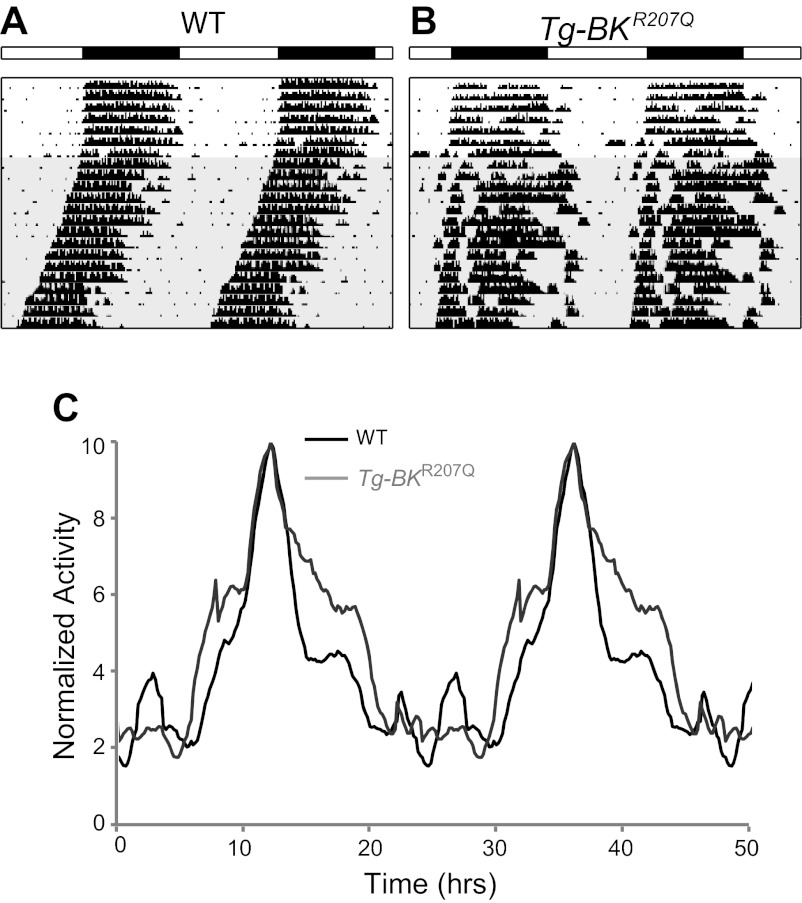
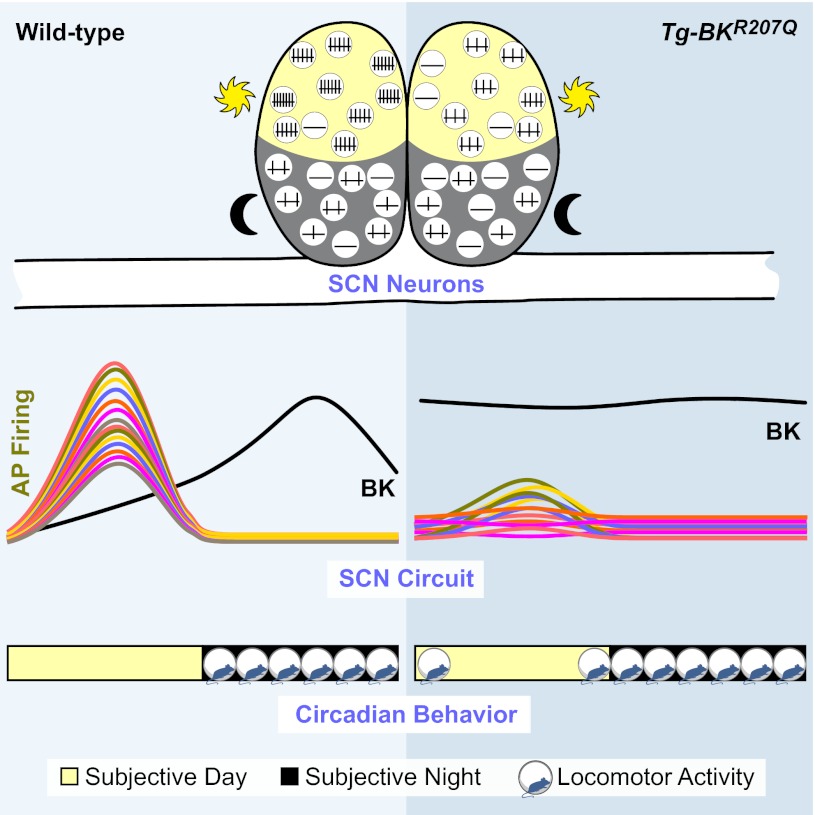
In mammals, almost all aspects of circadian rhythmicity are attributed to activity in a discrete neural circuit of the hypothalamus, the suprachiasmatic nucleus (SCN). A 24-h rhythm in spontaneous firing is the fundamental neural intermediary to circadian behavior, but the ionic mechanisms that pattern circuit rhythmicity, and the integrated impact on behavior, are not well studied. Here, we demonstrate that daily modulation of a major component of the nighttime-phased suppressive K(+) current, encoded by the BK Ca(2+)-activated K(+) current channel (K(Ca)1.1 or Kcnma1), is a critical arbiter of circadian rhythmicity in the SCN circuit. Aberrant induction of BK current during the day in transgenic mice using a Per1 promoter (Tg-BK(R207Q)) reduced SCN firing or silenced neurons, decreasing the circadian amplitude of the ensemble circuit rhythm. Changes in cellular and circuit excitability in Tg-BK(R207Q) SCNs were correlated with elongated behavioral active periods and enhanced responses to phase-shifting stimuli. Unexpectedly, despite the severe reduction in circuit amplitude, circadian behavioral amplitudes in Tg-BK(R207Q) mice were relatively normal. These data demonstrate that downregulation of the BK current during the day is essential for the high amplitude neural activity pattern in the SCN that restricts locomotor activity to the appropriate phase and maintains the clock's robustness against perturbation. However, a residually rhythmic subset prevails over the ensemble circuit to drive the fundamental circadian behavioral rhythm.
Figures
Similar articles
-
BK channels regulate spontaneous action potential rhythmicity in the suprachiasmatic nucleus.PLoS One. 2008;3(12):e3884. doi: 10.1371/journal.pone.0003884. Epub 2008 Dec 8. PLoS One. 2008. PMID: 19060951 Free PMC article.
-
Daily rhythmicity of large-conductance Ca2+ -activated K+ currents in suprachiasmatic nucleus neurons.Brain Res. 2006 Feb 3;1071(1):54-62. doi: 10.1016/j.brainres.2005.11.078. Epub 2006 Jan 17. Brain Res. 2006. PMID: 16412396
-
BK calcium-activated potassium channels regulate circadian behavioral rhythms and pacemaker output.Nat Neurosci. 2006 Aug;9(8):1041-9. doi: 10.1038/nn1740. Epub 2006 Jul 16. Nat Neurosci. 2006. PMID: 16845385 Free PMC article.
-
Ion Channels Controlling Circadian Rhythms in Suprachiasmatic Nucleus Excitability.Physiol Rev. 2020 Oct 1;100(4):1415-1454. doi: 10.1152/physrev.00027.2019. Epub 2020 Mar 12. Physiol Rev. 2020. PMID: 32163720 Free PMC article. Review.
-
Linking neural activity and molecular oscillations in the SCN.Nat Rev Neurosci. 2011 Sep 2;12(10):553-69. doi: 10.1038/nrn3086. Nat Rev Neurosci. 2011. PMID: 21886186 Free PMC article. Review.
Cited by
-
The circadian clock: A tale of genetic-electrical interplay and synaptic integration.Curr Opin Physiol. 2018 Oct;5:75-79. doi: 10.1016/j.cophys.2018.08.002. Epub 2018 Aug 24. Curr Opin Physiol. 2018. PMID: 31011692 Free PMC article.
-
Kv12-encoded K+ channels drive the day-night switch in the repetitive firing rates of SCN neurons.J Gen Physiol. 2023 Sep 4;155(9):e202213310. doi: 10.1085/jgp.202213310. Epub 2023 Jul 26. J Gen Physiol. 2023. PMID: 37516908 Free PMC article.
-
Membrane Currents, Gene Expression, and Circadian Clocks.Cold Spring Harb Perspect Biol. 2017 May 1;9(5):a027714. doi: 10.1101/cshperspect.a027714. Cold Spring Harb Perspect Biol. 2017. PMID: 28246182 Free PMC article. Review.
-
Circadian Control of Sleep by Melatonin via MT1-Dependent Activation of BK Channels in the Suprachiasmatic Nucleus.bioRxiv [Preprint]. 2025 May 23:2025.03.12.642893. doi: 10.1101/2025.03.12.642893. bioRxiv. 2025. PMID: 40475449 Free PMC article. Preprint.
-
Acetaldehyde-ethanol interactions on calcium-activated potassium (BK) channels in pituitary tumor (GH3) cells.Front Behav Neurosci. 2013 Jun 14;7:58. doi: 10.3389/fnbeh.2013.00058. eCollection 2013. Front Behav Neurosci. 2013. PMID: 23785316 Free PMC article.
References
-
- Abrahamson EE, Moore RY. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res 916: 172–191, 2001 - PubMed
-
- Akiyama M, Kouzu Y, Takahashi S, Wakamatsu H, Moriya T, Maetani M, Watanabe S, Tei H, Sakaki Y, Shibata S. Inhibition of light- or glutamate-induced mPer1 expression represses the phase shifts into the mouse circadian locomotor and suprachiasmatic firing rhythms. J Neurosci 19: 1115–1121, 1999 - PMC - PubMed
-
- Albus H, Bonnefont X, Chaves I, Yasui A, Doczy J, van der Horst GT, Meijer JH. Cryptochrome-deficient mice lack circadian electrical activity in the suprachiasmatic nuclei. Curr Biol 12: 1130–1133, 2002 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous