

Anti-IFN-α/β receptor antibody treatment ameliorates disease in lupus-predisposed mice
- PMID: 23175700
- PMCID: PMC3530925
- DOI: 10.4049/jimmunol.1201477
Anti-IFN-α/β receptor antibody treatment ameliorates disease in lupus-predisposed mice
Abstract
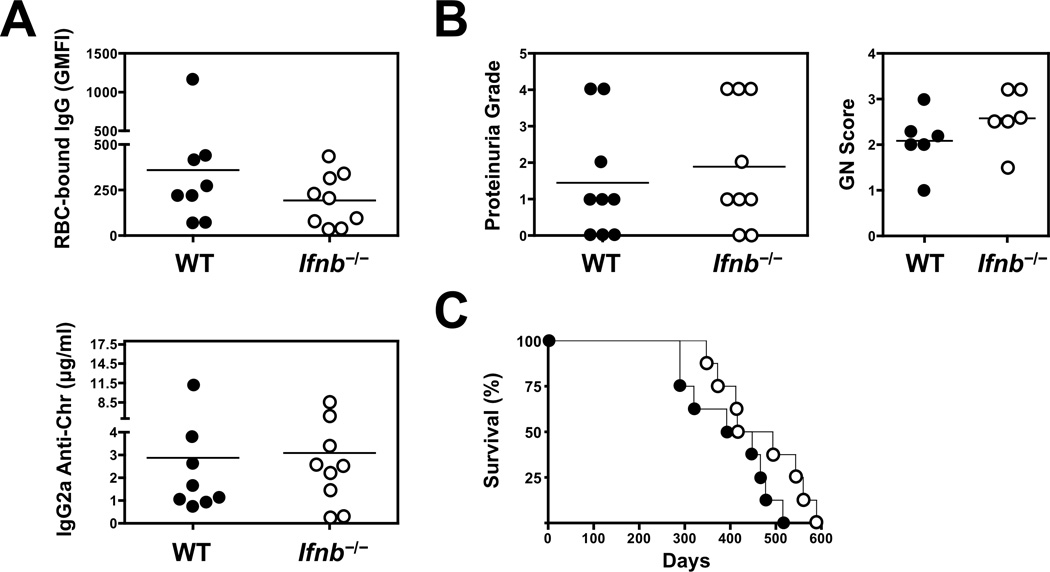
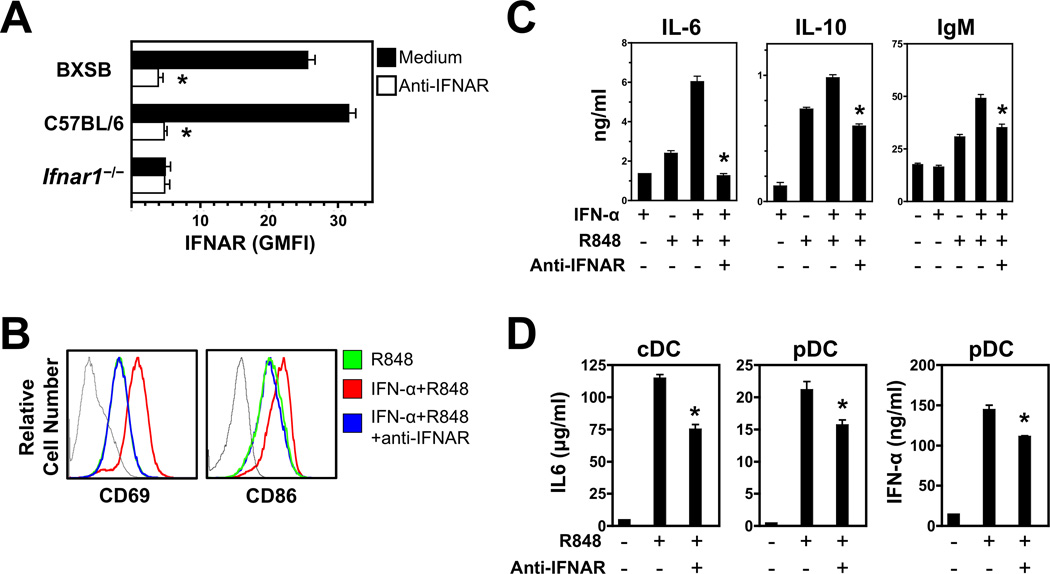
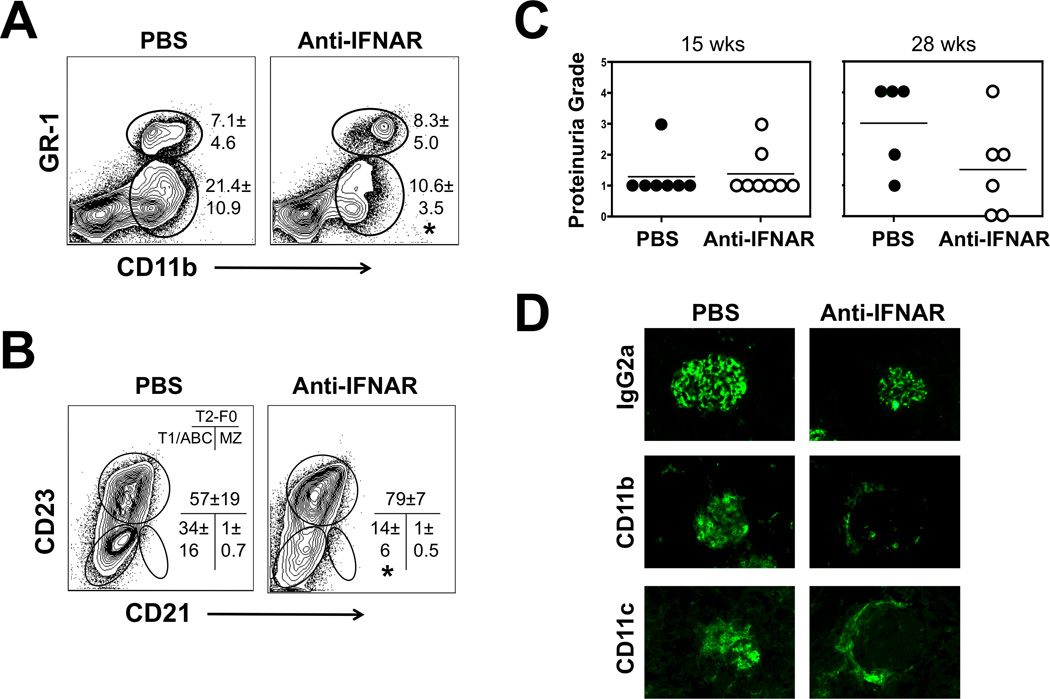
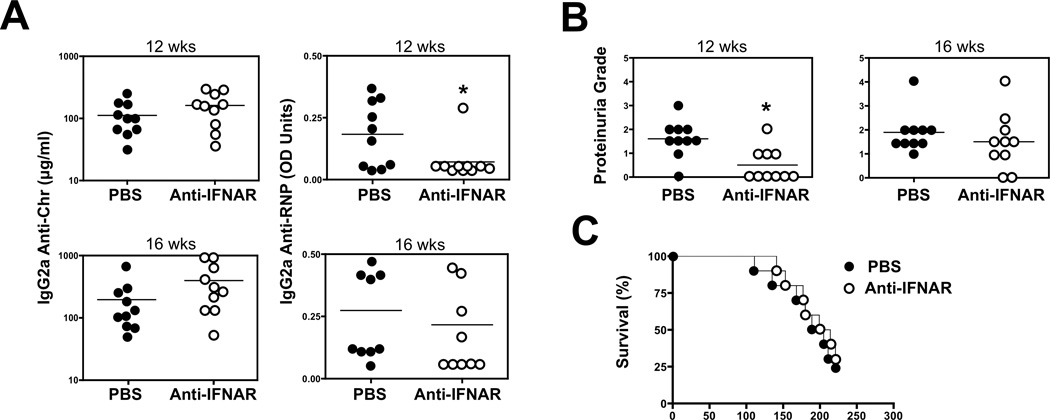
The demonstration in humans and mice that nucleic acid-sensing TLRs and type I IFNs are essential disease mediators is a milestone in delineating the mechanisms of lupus pathogenesis. In this study, we show that Ifnb gene deletion does not modify disease progression in NZB mice, thereby strongly implicating IFN-α subtypes as the principal pathogenic effectors. We further document that long-term treatment of male BXSB mice with an anti-IFN-α/β receptor Ab of mouse origin reduced serologic, cellular, and histologic disease manifestations and extended survival, suggesting that disease acceleration by the Tlr7 gene duplication in this model is mediated by type I IFN signaling. The efficacy of this treatment in BXSB mice was clearly evident when applied early in the disease process, but only partial reductions in some disease characteristics were observed when treatment was initiated at later stages. A transient therapeutic effect was also noted in the MRL-Fas(lpr) model, although overall mortality was unaffected. The combined findings suggest that IFN-α/β receptor blockade, particularly when started at early disease stages, may be a useful treatment approach for human systemic lupus erythematosus and other autoimmune syndromes.
Figures



References
-
- Theofilopoulos AN, Baccala R, Beutler B, Kono DH. Type I interferons (alpha/beta) in immunity and autoimmunity. Annu Rev Immunol. 2005;23:307–335. - PubMed
-
- Banchereau J, Pascual V. Type I interferon in systemic lupus erythematosus and other autoimmune diseases. Immunity. 2006;25:383–392. - PubMed
-
- Ronnblom L, Eloranta ML, Alm GV. The type I interferon system in systemic lupus erythematosus. Arthritis Rheum. 2006;54:408–420. - PubMed
-
- Marshak-Rothstein A, Rifkin IR. Immunologically active autoantigens: the role of toll-like receptors in the development of chronic inflammatory disease. Annu Rev Immunol. 2007;25:419–441. - PubMed
-
- Baccala R, Hoebe K, Kono DH, Beutler B, Theofilopoulos AN. TLR-dependent and TLR-independent pathways of type I interferon induction in systemic autoimmunity. Nat Med. 2007;13:543–551. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
