

KAYAK-α modulates circadian transcriptional feedback loops in Drosophila pacemaker neurons
- PMID: 23175847
- PMCID: PMC3528402
- DOI: 10.1523/JNEUROSCI.1888-12.2012
KAYAK-α modulates circadian transcriptional feedback loops in Drosophila pacemaker neurons
Abstract
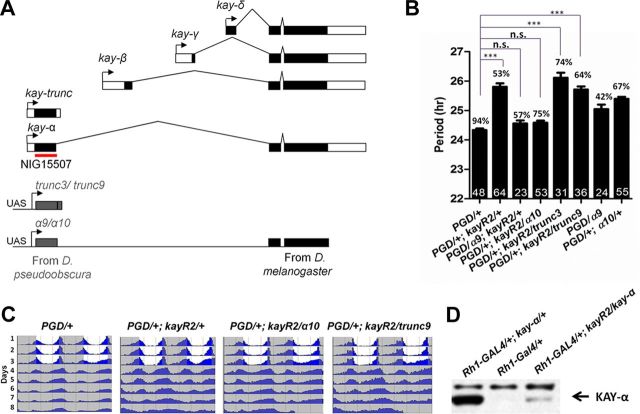
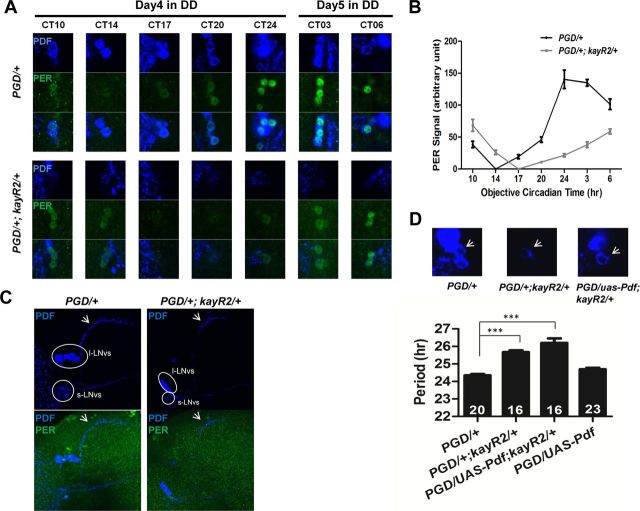
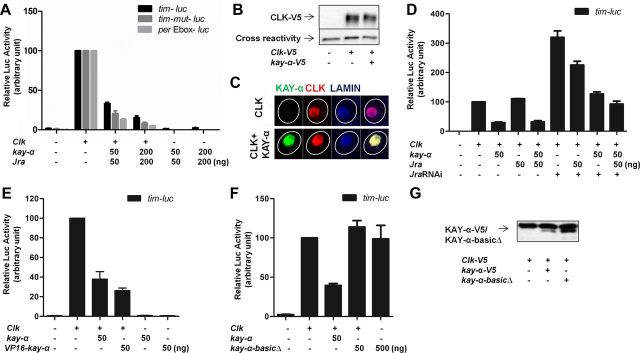
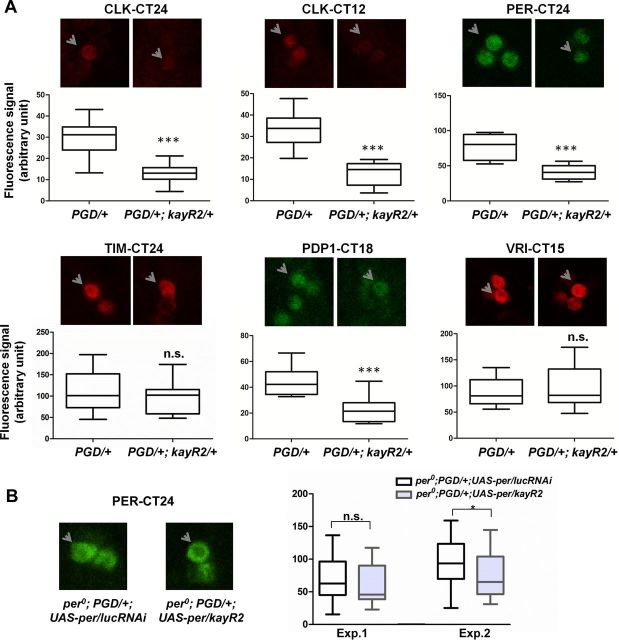
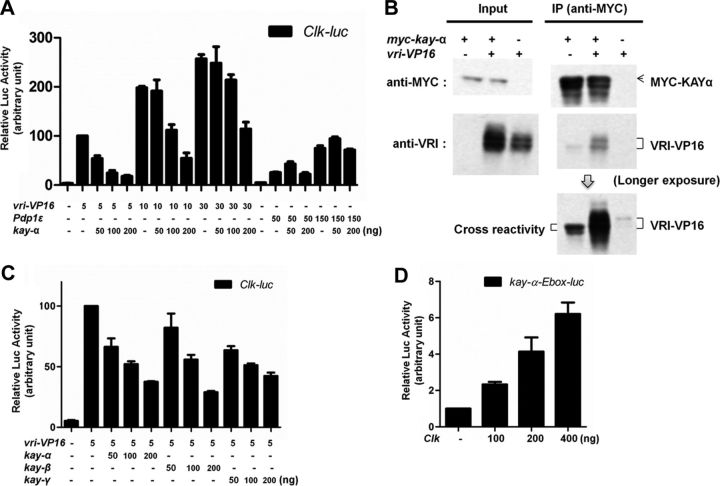
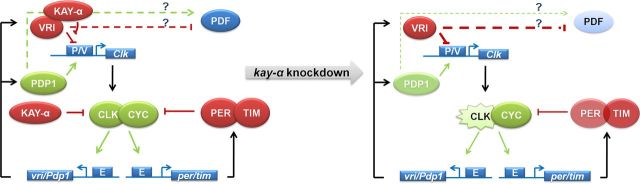
Circadian rhythms are generated by well-conserved interlocked transcriptional feedback loops in animals. In Drosophila, the dimeric transcription factor CLOCK/CYCLE (CLK/CYC) promotes period (per), timeless (tim), vrille (vri), and PAR-domain protein 1 (Pdp1) transcription. PER and TIM negatively feed back on CLK/CYC transcriptional activity, whereas VRI and PDP1 negatively and positively regulate Clk transcription, respectively. Here, we show that the α isoform of the Drosophila FOS homolog KAYAK (KAY) is required for normal circadian behavior. KAY-α downregulation in circadian pacemaker neurons increases period length by 1.5 h. This behavioral phenotype is correlated with decreased expression of several circadian proteins. The strongest effects are on CLK and the neuropeptide PIGMENT DISPERSING FACTOR, which are both under VRI and PDP1 control. Consistently, KAY-α can bind to VRI and inhibit its interaction with the Clk promoter. Interestingly, KAY-α can also repress CLK activity. Hence, in flies with low KAY-α levels, CLK derepression would partially compensate for increased VRI repression, thus attenuating the consequences of KAY-α downregulation on CLK targets. We propose that the double role of KAY-α in the two transcriptional loops controlling Drosophila circadian behavior brings precision and stability to their oscillations.
Figures

Similar articles
-
VRILLE Controls PDF Neuropeptide Accumulation and Arborization Rhythms in Small Ventrolateral Neurons to Drive Rhythmic Behavior in Drosophila.Curr Biol. 2017 Nov 20;27(22):3442-3453.e4. doi: 10.1016/j.cub.2017.10.010. Epub 2017 Nov 2. Curr Biol. 2017. PMID: 29103936
-
The Drosophila Receptor Protein Tyrosine Phosphatase LAR Is Required for Development of Circadian Pacemaker Neuron Processes That Support Rhythmic Activity in Constant Darkness But Not during Light/Dark Cycles.J Neurosci. 2016 Mar 30;36(13):3860-70. doi: 10.1523/JNEUROSCI.4523-15.2016. J Neurosci. 2016. PMID: 27030770 Free PMC article.
-
PDP1epsilon functions downstream of the circadian oscillator to mediate behavioral rhythms.J Neurosci. 2007 Mar 7;27(10):2539-47. doi: 10.1523/JNEUROSCI.4870-06.2007. J Neurosci. 2007. PMID: 17344391 Free PMC article.
-
Transcriptional feedback loop regulation, function, and ontogeny in Drosophila.Cold Spring Harb Symp Quant Biol. 2007;72:437-44. doi: 10.1101/sqb.2007.72.009. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419302 Free PMC article. Review.
-
Regulating a circadian clock's period, phase and amplitude by phosphorylation: insights from Drosophila.J Biochem. 2006 Nov;140(5):609-17. doi: 10.1093/jb/mvj198. Epub 2006 Sep 29. J Biochem. 2006. PMID: 17012288 Review.
Cited by
-
Polycomb regulates circadian rhythms in Drosophila in clock neurons.Life Sci Alliance. 2023 Nov 1;7(1):e202302140. doi: 10.26508/lsa.202302140. Print 2024 Jan. Life Sci Alliance. 2023. PMID: 37914396 Free PMC article.
-
Integrated omics in Drosophila uncover a circadian kinome.Nat Commun. 2020 Jun 1;11(1):2710. doi: 10.1038/s41467-020-16514-z. Nat Commun. 2020. PMID: 32483184 Free PMC article.
-
Evolutionary divergence of core and post-translational circadian clock genes in the pitcher-plant mosquito, Wyeomyia smithii.BMC Genomics. 2015 Oct 6;16:754. doi: 10.1186/s12864-015-1937-y. BMC Genomics. 2015. PMID: 26444857 Free PMC article.
-
Selection for long and short sleep duration in Drosophila melanogaster reveals the complex genetic network underlying natural variation in sleep.PLoS Genet. 2017 Dec 14;13(12):e1007098. doi: 10.1371/journal.pgen.1007098. eCollection 2017 Dec. PLoS Genet. 2017. PMID: 29240764 Free PMC article.
-
Circadian period integrates network information through activation of the BMP signaling pathway.PLoS Biol. 2013 Dec;11(12):e1001733. doi: 10.1371/journal.pbio.1001733. Epub 2013 Dec 10. PLoS Biol. 2013. PMID: 24339749 Free PMC article.
References
-
- Akten B, Jauch E, Genova GK, Kim EY, Edery I, Raabe T, Jackson FR. A role for CK2 in the Drosophila circadian oscillator. Nat Neurosci. 2003;6:251–257. - PubMed
-
- Blanchardon E, Grima B, Klarsfeld A, Chélot E, Hardin PE, Préat T, Rouyer F. Defining the role of Drosophila lateral neurons in the control of circadian rhythms in motor activity and eclosion by targeted genetic ablation and PERIOD protein overexpression. Eur J Neurosci. 2001;13:871–888. - PubMed
-
- Blau J, Young MW. Cycling vrille expression is required for a functional Drosophila clock. Cell. 1999;99:661–671. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials