

Stroma-derived interleukin-34 controls the development and maintenance of langerhans cells and the maintenance of microglia
- PMID: 23177320
- PMCID: PMC4291117
- DOI: 10.1016/j.immuni.2012.11.001
Stroma-derived interleukin-34 controls the development and maintenance of langerhans cells and the maintenance of microglia
Abstract
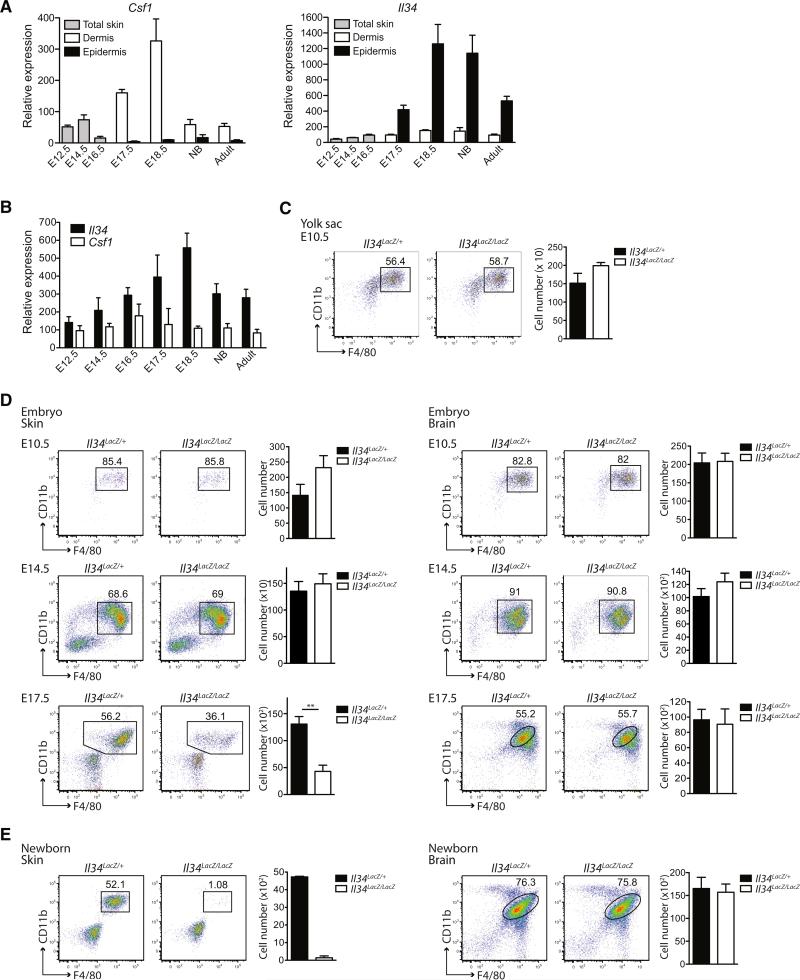
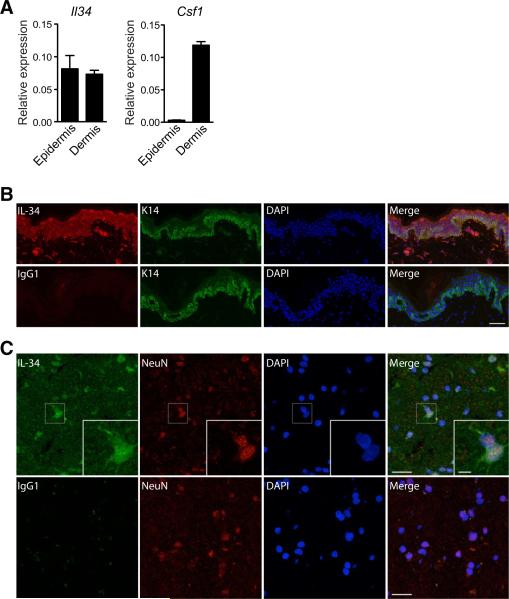
Colony stimulating factor-1 (Csf-1) receptor and its ligand Csf-1 control macrophage development, maintenance, and function. The development of both Langerhans cells (LCs) and microglia is highly dependent on Csf-1 receptor signaling but independent of Csf-1. Here we show that in both mice and humans, interleukin-34 (IL-34), an alternative ligand for Csf-1 receptor, is produced by keratinocytes in the epidermis and by neurons in the brain. Mice lacking IL-34 displayed a marked reduction of LCs and a decrease of microglia, whereas monocytes, dermal, and lymphoid tissue macrophages and DCs were unaffected. We identified IL-34 as a nonredundant cytokine for the development of LCs during embryogenesis as well as for their homeostasis in the adult skin. Whereas inflammation-induced repopulation of LCs appears to be dependent on Csf-1, once inflammation is resolved, LC survival is again IL-34-dependent. In contrast, microglia and their yolk sac precursors develop independently of IL-34 but rely on it for their maintenance in the adult brain.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia.Nat Immunol. 2012 Jun 24;13(8):753-60. doi: 10.1038/ni.2360. Nat Immunol. 2012. PMID: 22729249 Free PMC article.
-
Nonredundant roles of keratinocyte-derived IL-34 and neutrophil-derived CSF1 in Langerhans cell renewal in the steady state and during inflammation.Eur J Immunol. 2016 Mar;46(3):552-9. doi: 10.1002/eji.201545917. Epub 2015 Dec 28. Eur J Immunol. 2016. PMID: 26634935 Free PMC article.
-
Interkeukin-34, a cytokine crucial for the differentiation and maintenance of tissue resident macrophages and Langerhans cells.Eur J Immunol. 2014 Jun;44(6):1575-81. doi: 10.1002/eji.201344365. Epub 2014 May 22. Eur J Immunol. 2014. PMID: 24737461 Free PMC article. Review.
-
IL-34 and CSF-1: similarities and differences.J Bone Miner Metab. 2013 Sep;31(5):486-95. doi: 10.1007/s00774-013-0476-3. Epub 2013 Jun 6. J Bone Miner Metab. 2013. PMID: 23740288 Review.
-
IL-23 from Langerhans cells is required for the development of imiquimod-induced psoriasis-like dermatitis by induction of IL-17A-producing γδ T cells.J Invest Dermatol. 2014 Jul;134(7):1912-1921. doi: 10.1038/jid.2014.98. Epub 2014 Feb 25. J Invest Dermatol. 2014. PMID: 24569709
Cited by
-
Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage.J Leukoc Biol. 2016 Sep;100(3):481-9. doi: 10.1189/jlb.3RU0316-144R. Epub 2016 Jun 28. J Leukoc Biol. 2016. PMID: 27354413 Free PMC article. Review.
-
IL-34 attenuates acute T cell-mediated rejection following renal transplantation by upregulating M2 macrophages polarization.Heliyon. 2024 Jan 3;10(1):e24028. doi: 10.1016/j.heliyon.2024.e24028. eCollection 2024 Jan 15. Heliyon. 2024. PMID: 38230243 Free PMC article.
-
Emerging Roles for CSF-1 Receptor and its Ligands in the Nervous System.Trends Neurosci. 2016 Jun;39(6):378-393. doi: 10.1016/j.tins.2016.03.005. Epub 2016 Apr 12. Trends Neurosci. 2016. PMID: 27083478 Free PMC article. Review.
-
RARα supports the development of Langerhans cells and langerin-expressing conventional dendritic cells.Nat Commun. 2018 Sep 25;9(1):3896. doi: 10.1038/s41467-018-06341-8. Nat Commun. 2018. PMID: 30254197 Free PMC article.
-
Alveolar macrophages develop from fetal monocytes that differentiate into long-lived cells in the first week of life via GM-CSF.J Exp Med. 2013 Sep 23;210(10):1977-92. doi: 10.1084/jem.20131199. Epub 2013 Sep 16. J Exp Med. 2013. PMID: 24043763 Free PMC article.
References
-
- Chihara T, Suzu S, Hassan R, Chutiwitoonchai N, Hiyoshi M, Motoyoshi K, Kimura F, Okada S. IL-34 and M-CSF share the receptor Fms but are not identical in biological activity and signal activation. Cell Death Differ. 2010;17:1917–1927. - PubMed
-
- Dai XM, Ryan GR, Hapel AJ, Dominguez MG, Russell RG, Kapp S, Sylvestre V, Stanley ER. Targeted disruption of the mouse colony-stimulating factor 1 receptor gene results in osteopetrosis, mononuclear phagocyte deficiency, increased primitive progenitor cell frequencies, and reproductive defects. Blood. 2002;99:111–120. - PubMed
-
- Garceau V, Smith J, Paton IR, Davey M, Fares MA, Sester DP, Burt DW, Hume DA. Pivotal Advance: Avian colony-stimulating factor 1 (CSF-1), interleukin-34 (IL-34), and CSF-1 receptor genes and gene products. J. Leukoc. Biol. 2010;87:753–764. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous