

Anterior-posterior direction opponency in the superficial mouse lateral geniculate nucleus
- PMID: 23177957
- PMCID: PMC3517882
- DOI: 10.1016/j.neuron.2012.09.021
Anterior-posterior direction opponency in the superficial mouse lateral geniculate nucleus
Abstract
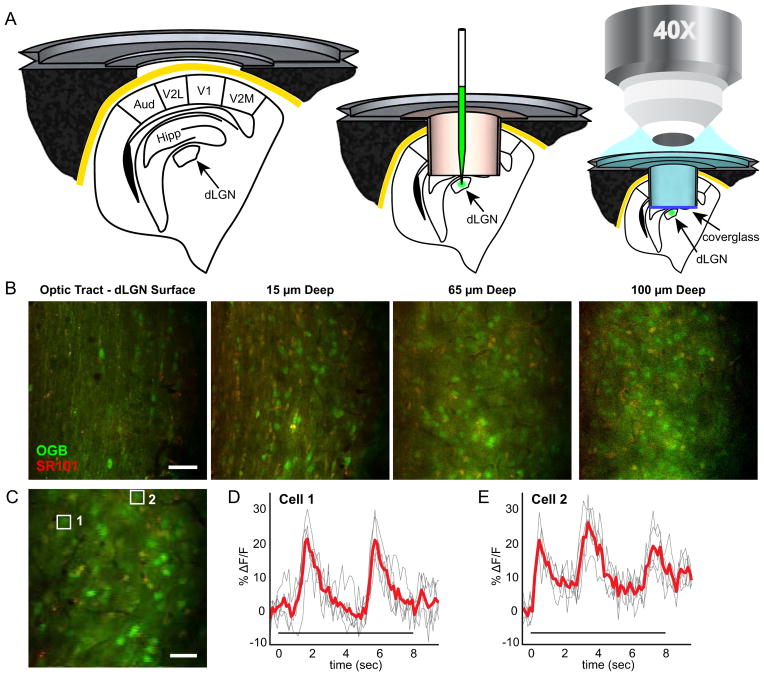
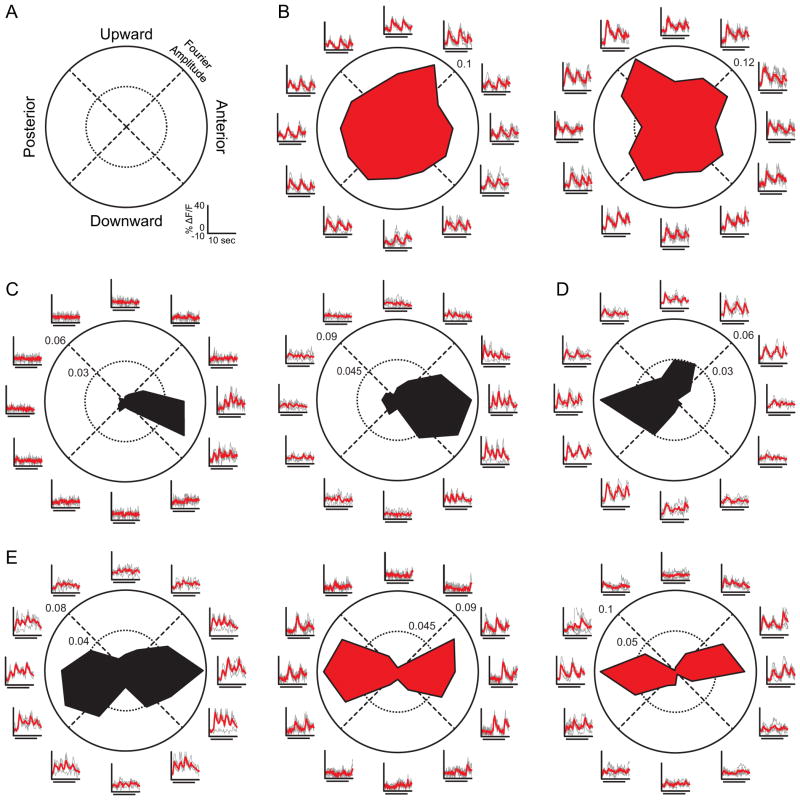
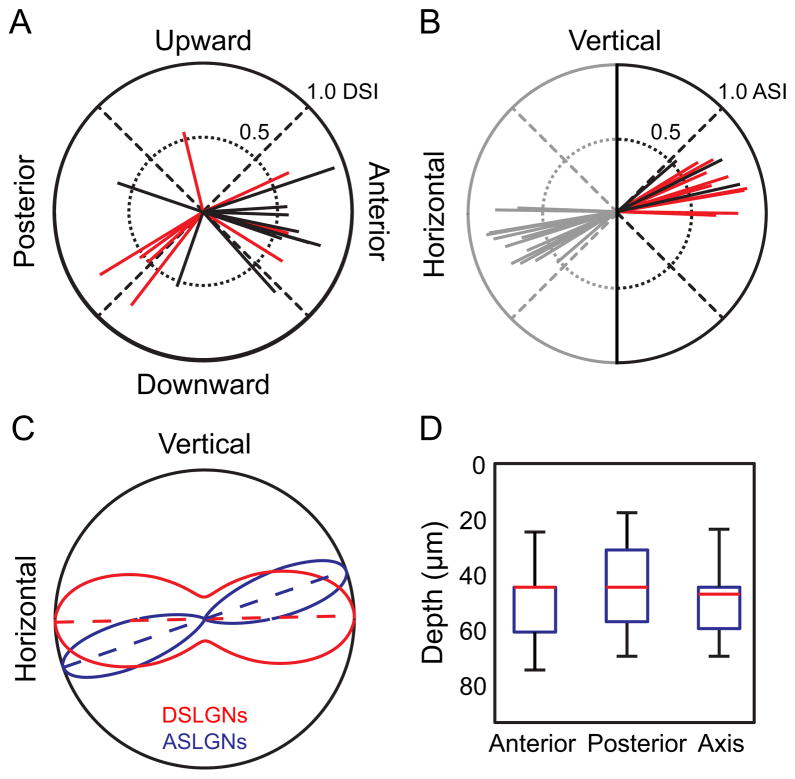
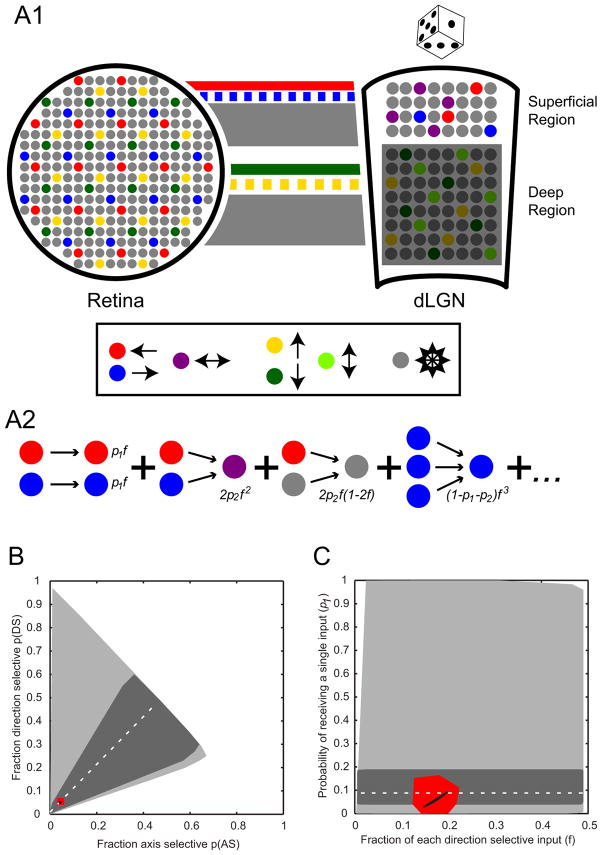
We show functional-anatomical organization of motion direction in mouse dorsal lateral geniculate nucleus (dLGN) using two-photon calcium imaging of dense populations in thalamus. Surprisingly, the superficial 75 μm region contains anterior and posterior direction-selective neurons (DSLGNs) intermingled with nondirection-selective neurons, while upward- and downward-selective neurons are nearly absent. Unexpectedly, the remaining neurons encode both anterior and posterior directions, forming horizontal motion-axis selectivity. A model of random wiring consistent with these results makes quantitative predictions about the connectivity of direction-selective retinal ganglion cell (DSRGC) inputs to the superficial dLGN. DSLGNs are more sharply tuned than DSRGCs. These results suggest that dLGN maintains and sharpens retinal direction selectivity and integrates opposing DSRGC subtypes in a functional-anatomical region, perhaps forming a feature representation for horizontal-axis motion, contrary to dLGN being a simple relay. Furthermore, they support recent conjecture that cortical direction and orientation selectivity emerge in part from a previously undescribed motion-selective retinogeniculate pathway.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Briggman KL, Helmstaedter M, Denk W. Wiring specificity in the direction-selectivity circuit of the retina. Nature. 2011;471:183–188. - PubMed
-
- Chen C, Regehr WG. Developmental remodeling of the retinogeniculate synapse. Neuron. 2000;28:955–966. - PubMed
-
- Cleland BG, Dubin MW, Levick WR. Simultaneous recording of input and output of lateral geniculate neurones. Nature New Biol. 1971a;231:191–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
