

A fructose receptor functions as a nutrient sensor in the Drosophila brain
- PMID: 23178127
- PMCID: PMC3509419
- DOI: 10.1016/j.cell.2012.10.024
A fructose receptor functions as a nutrient sensor in the Drosophila brain
Abstract
Internal nutrient sensors play important roles in feeding behavior, yet their molecular structure and mechanism of action are poorly understood. Using Ca(2+) imaging and behavioral assays, we show that the gustatory receptor 43a (Gr43a) functions as a narrowly tuned fructose receptor in taste neurons. Remarkably, Gr43a also functions as a fructose receptor in the brain. Interestingly, hemolymph fructose levels are tightly linked to feeding status: after nutritious carbohydrate consumption, fructose levels rise several fold and reach a concentration sufficient to activate Gr43a in the brain. By using different feeding paradigms and artificial activation of Gr43a-expressing brain neurons, we show that Gr43a is both necessary and sufficient to sense hemolymph fructose and promote feeding in hungry flies but suppress feeding in satiated flies. Thus, our studies indicate that the Gr43a-expressing brain neurons function as a nutrient sensor for hemolymph fructose and assign opposing valence to feeding experiences in a satiation-dependent manner.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
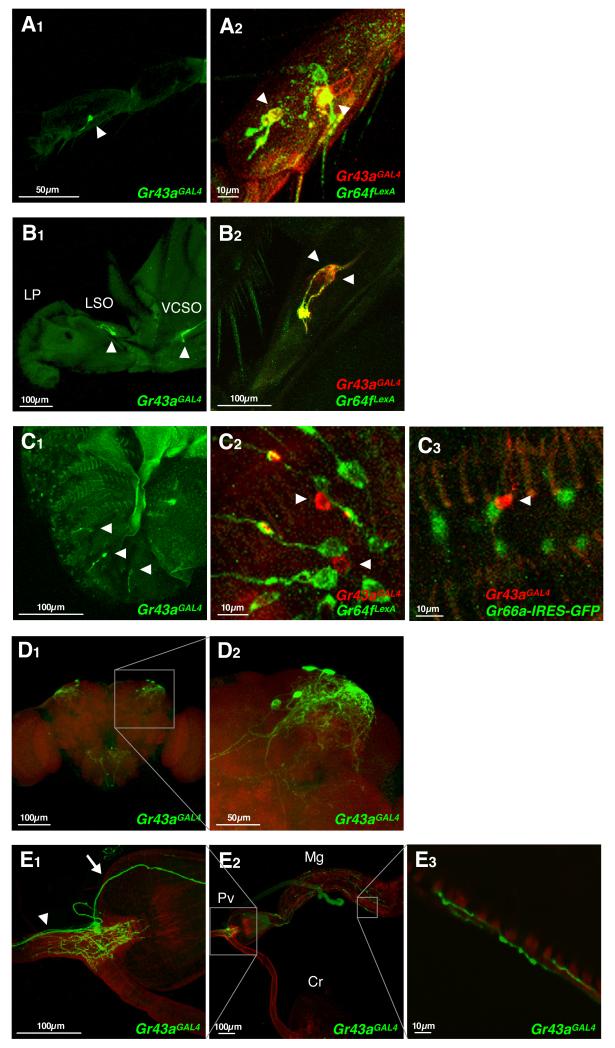
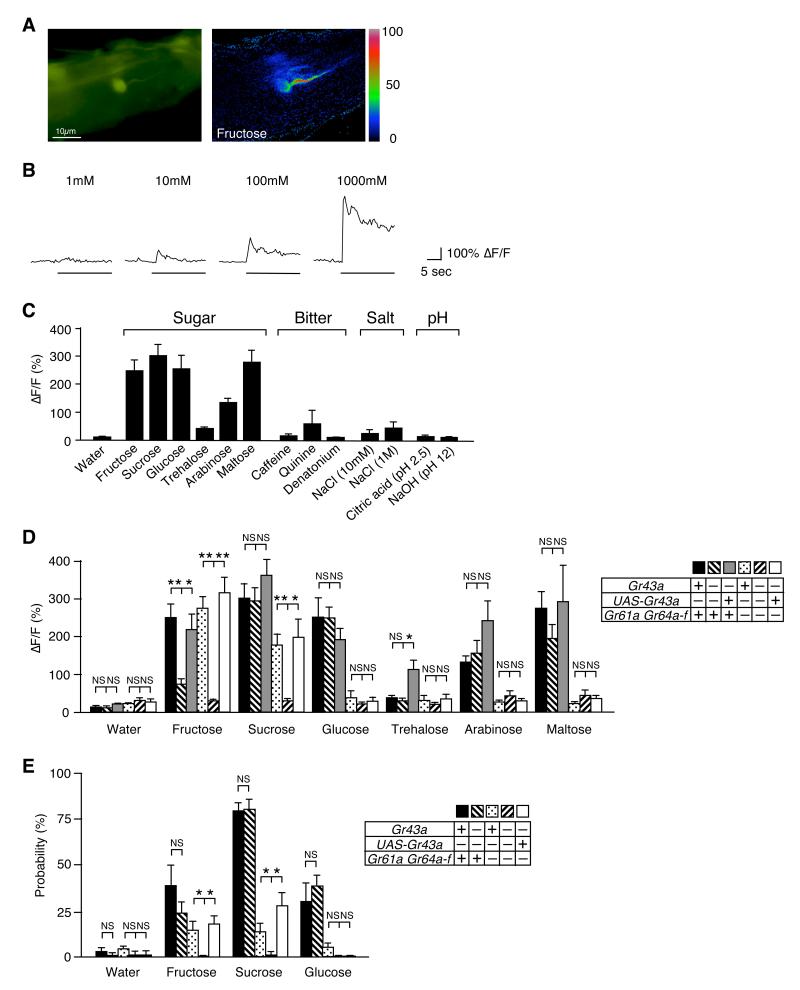
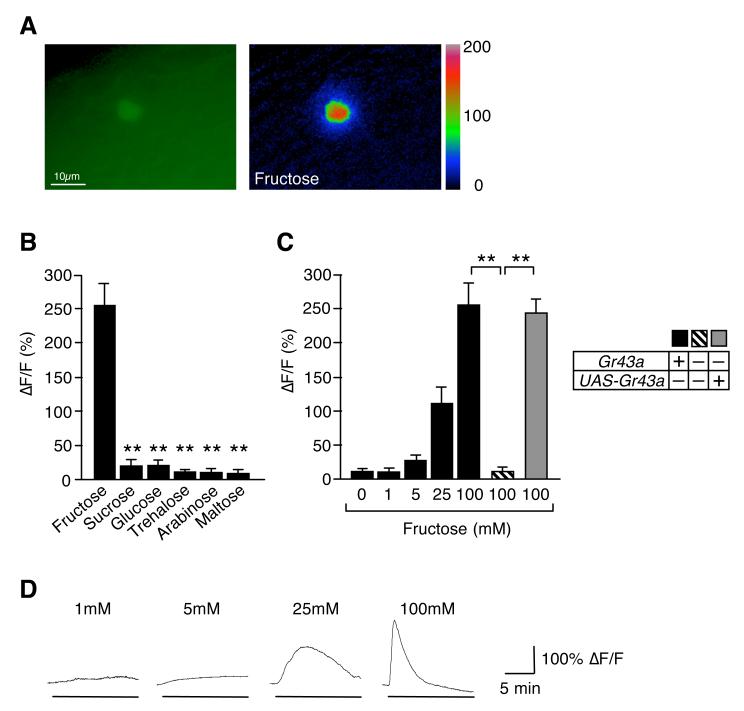
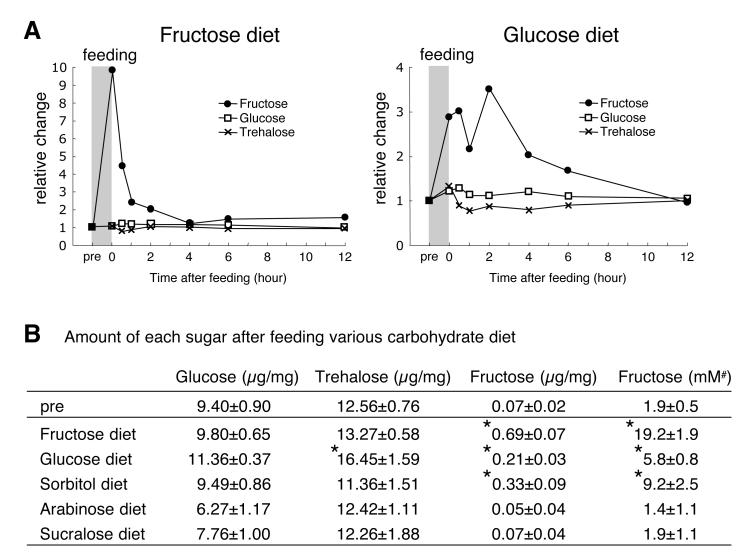
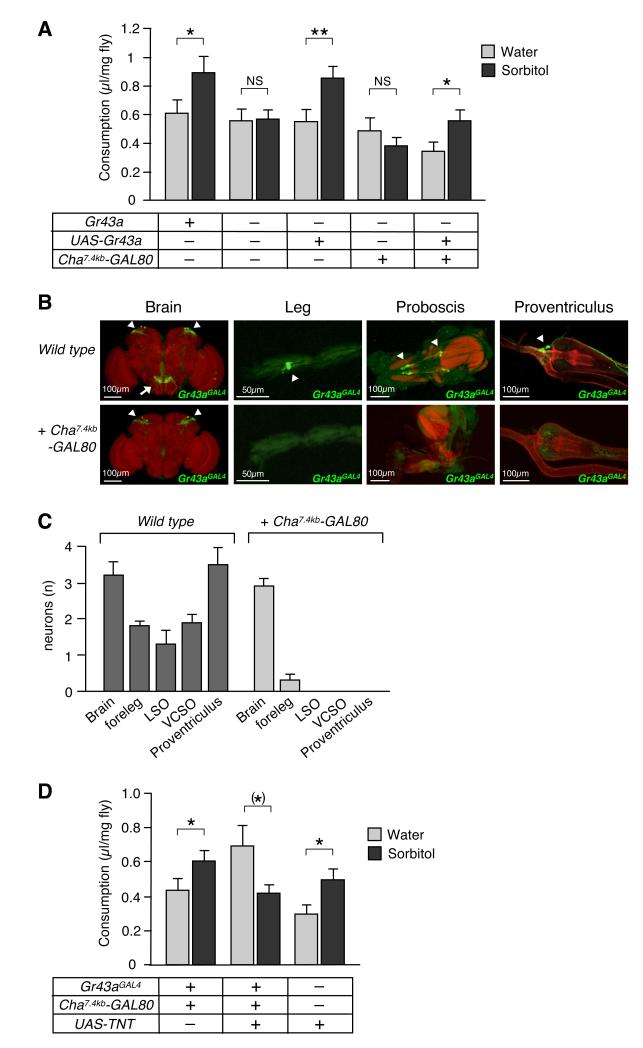
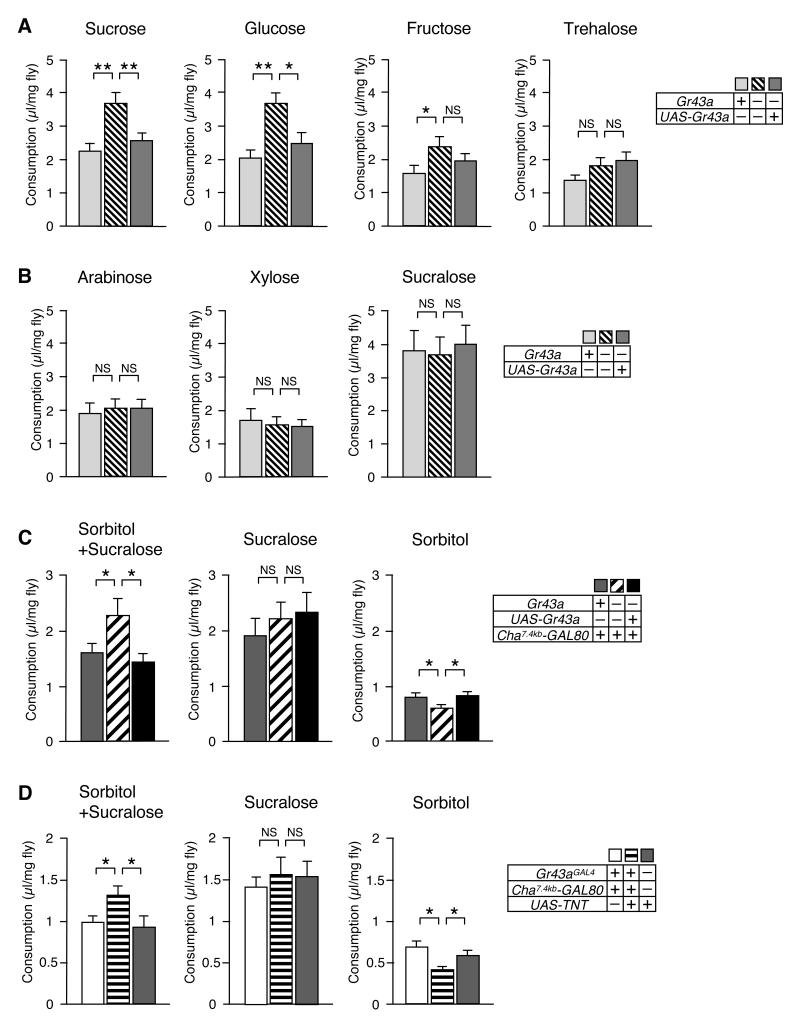
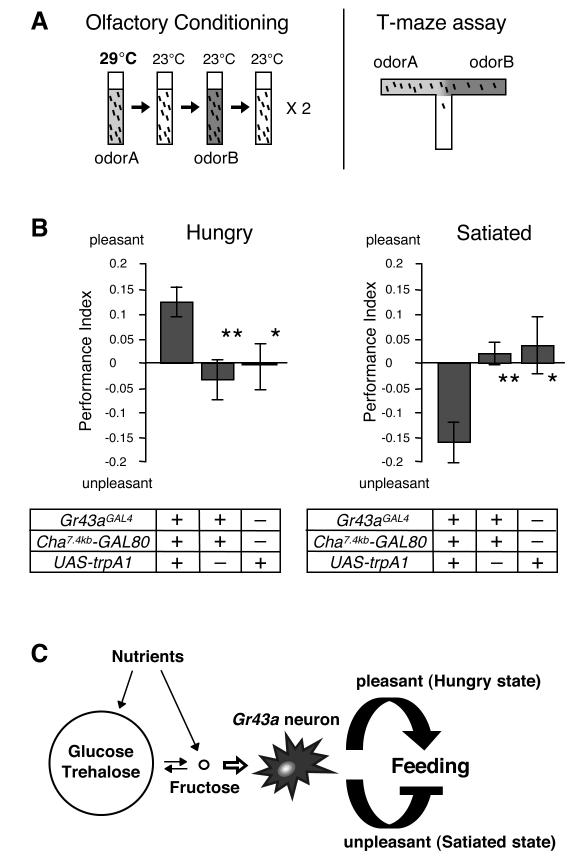
Comment in
-
Let the sensor fit the diet.Cell Metab. 2012 Dec 5;16(6):689-90. doi: 10.1016/j.cmet.2012.11.008. Cell Metab. 2012. PMID: 23217252 Free PMC article.
References
-
- Buch S, Melcher C, Bauer M, Katzenberger J, Pankratz MJ. Opposing effects of dietary protein and sugar regulate a transcriptional target of Drosophila insulin-like peptide signaling. Cell Metab. 2008;7:321–332. - PubMed
-
- Chapman R. The Insects: Structure and Function. 4th edn Cambridge University Press; Cambridge, New York, Melbourne, Madrid, Cape Town: 1998.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
