

Presphenoidal synchondrosis fusion in DBA/2J mice
- PMID: 23179633
- PMCID: PMC3560942
- DOI: 10.1007/s00335-012-9437-8
Presphenoidal synchondrosis fusion in DBA/2J mice
Abstract
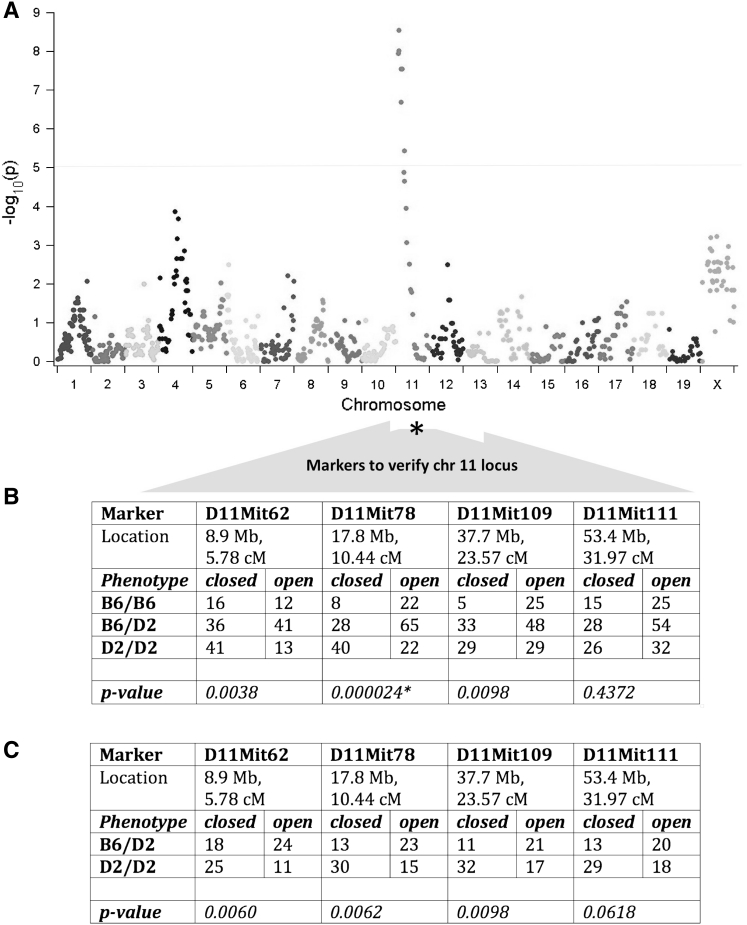
Cranial base growth plates are important centers of longitudinal growth in the skull and are responsible for the proper anterior placement of the face and the stimulation of normal cranial vault development. We report that the presphenoidal synchondrosis (PSS), a midline growth plate of the cranial base, closes in the DBA/2J mouse strain but not in other common inbred strains. We investigated the genetics of PSS closure in DBA/2J mice by evaluating F1, F1 backcross, and/or F1 intercross offspring from matings with C57BL/6J and DBA/1J mice, whose PSS remain open. We observed that PSS closure is genetically determined, but not inherited as a simple Mendelian trait. Employing a genome-wide SNP array, we identified a region on chromosome 11 in the C57BL/6J strain that affected the frequency of PSS closure in F1 backcross and F1 intercross offspring. The equivalent region in the DBA/1J strain did not affect PSS closure in F1 intercross offspring. We conclude that PSS closure in the DBA/2J strain is complex and modified by different loci when outcrossed with C57BL/6J and DBA/1J mice.
Figures


Similar articles
-
Sex-restricted non-Mendelian inheritance of mouse chromosome 11 in the offspring of crosses between C57BL/6J and (C57BL/6J x DBA/2J)F1 mice.Mamm Genome. 1998 Oct;9(10):812-5. doi: 10.1007/s003359900872. Mamm Genome. 1998. PMID: 9745035
-
Segregation analysis of the testis-determining autosomal trait, Tda, that differs between the C57Bl/6J and DBA/2J mouse strains suggests a multigenic threshold model.Genome. 1996 Apr;39(2):322-35. doi: 10.1139/g96-043. Genome. 1996. PMID: 8984004
-
Behavioural analysis of congenic mouse strains confirms stress-responsive Loci on chromosomes 1 and 12.Behav Genet. 2008 Jul;38(4):407-16. doi: 10.1007/s10519-008-9206-3. Epub 2008 Apr 1. Behav Genet. 2008. PMID: 18379869
-
The testis-determining autosomal trait, Tda-1, of C57BL/6J is determined by more than a single autosomal gene when compared with DBA/2J mice.Genome. 1994 Apr;37(2):296-304. doi: 10.1139/g94-041. Genome. 1994. PMID: 8200518
-
Use of recombinant inbred strains for studying genetic determinants of responses to alcohol.Alcohol Alcohol Suppl. 1994;2:67-71. Alcohol Alcohol Suppl. 1994. PMID: 8974318 Review.
Cited by
-
Postnatal Ontogeny of the Cranial Base and Craniofacial Skeleton in Male C57BL/6J Mice: A Reference Standard for Quantitative Analysis.Front Physiol. 2016 Jan 12;6:417. doi: 10.3389/fphys.2015.00417. eCollection 2015. Front Physiol. 2016. PMID: 26793119 Free PMC article.
-
Collagen mutation and age contribute to differential craniofacial phenotypes in mouse models of osteogenesis imperfecta.JBMR Plus. 2024 Jan 4;8(1):ziad004. doi: 10.1093/jbmrpl/ziad004. eCollection 2024 Jan. JBMR Plus. 2024. PMID: 38690127 Free PMC article.
References
-
- Bjork A. Cranial base development. Am J Orthod. 1955;41:199–255. doi: 10.1016/0002-9416(55)90005-1. - DOI
-
- Bourgeois P, Bolcato-Bellemin AL, Danse JM, Bloch-Zupan A, Yoshiba K, Stoetzel C, Perrin-Schmitt F. The variable expressivity and incomplete penetrance of the twist-null heterozygous mouse phenotype resemble those of human Saethre–Chotzen syndrome. Hum Mol Genet. 1998;7:945–957. doi: 10.1093/hmg/7.6.945. - DOI - PubMed
-
- Bowen ME, Boyden ED, Holm IA, Campos-Xavier B, Bonafe L, Superti-Furga A, Ikegawa S, Cormier-Daire V, Bovee JV, Pansuriya TC, de Sousa SB, Savarirayan R, Andreucci E, Vikkula M, Garavelli L, Pottinger C, Ogino T, Sakai A, Regazzoni BM, Wuyts W, Sangiorgi L, Pedrini E, Zhu M, Kozakewich HP, Kasser JR, Seidman JG, Kurek KC, Warman ML. Loss-of-function mutations in PTPN11 cause metachondromatosis, but not Ollier disease or Maffucci syndrome. PLoS Genet. 2011;7:e1002050. doi: 10.1371/journal.pgen.1002050. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
