

The spatial and temporal origin of chandelier cells in mouse neocortex
- PMID: 23180771
- PMCID: PMC4017638
- DOI: 10.1126/science.1227622
The spatial and temporal origin of chandelier cells in mouse neocortex
Abstract
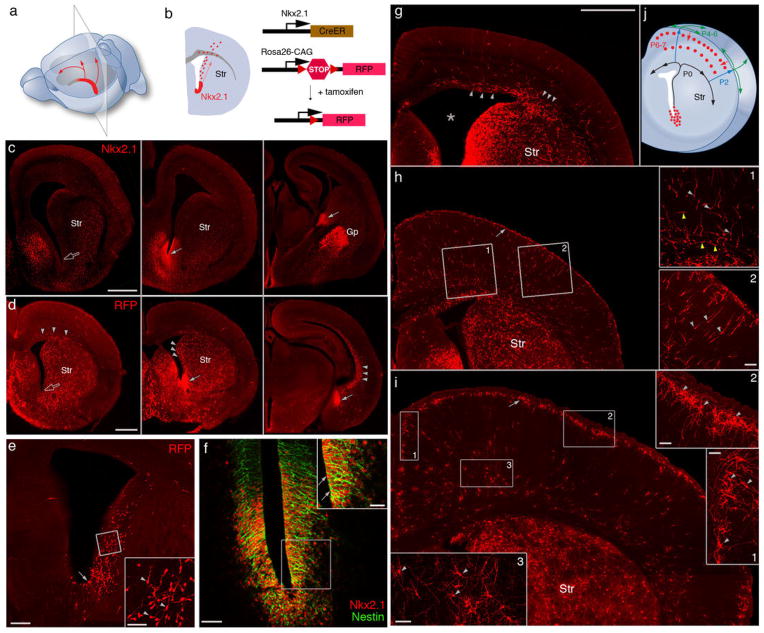
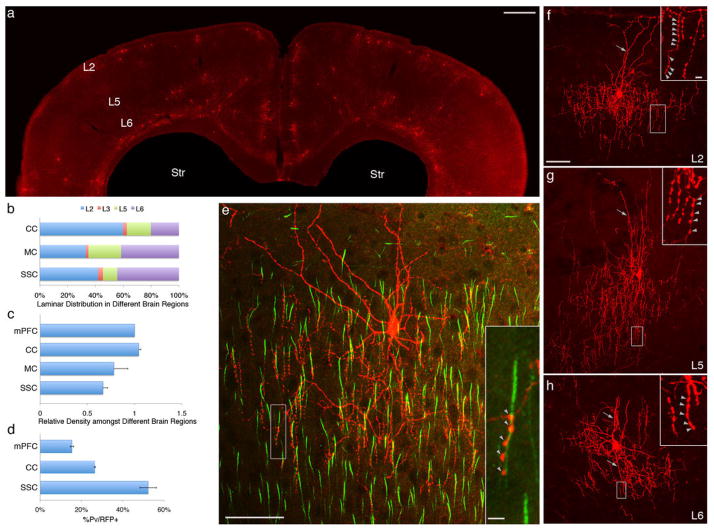
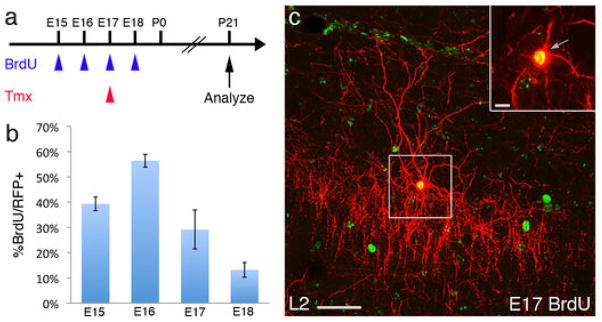
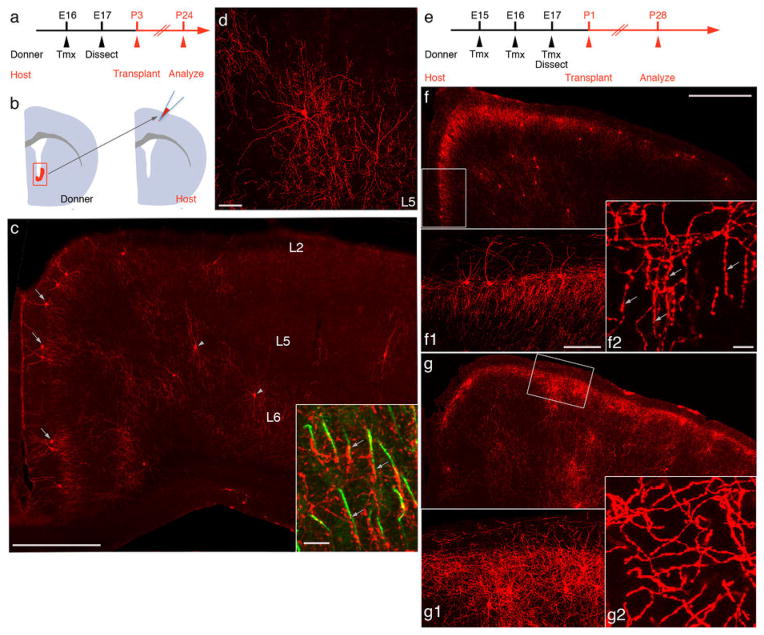
Diverse γ-aminobutyric acid-releasing interneurons regulate the functional organization of cortical circuits and derive from multiple embryonic sources. It remains unclear to what extent embryonic origin influences interneuron specification and cortical integration because of difficulties in tracking defined cell types. Here, we followed the developmental trajectory of chandelier cells (ChCs), the most distinct interneurons that innervate the axon initial segment of pyramidal neurons and control action potential initiation. ChCs mainly derive from the ventral germinal zone of the lateral ventricle during late gestation and require the homeodomain protein Nkx2.1 for their specification. They migrate with stereotyped routes and schedule and achieve specific laminar distribution in the cortex. The developmental specification of this bona fide interneuron type likely contributes to the assembly of a cortical circuit motif.
Figures
Comment in
-
Neuroscience. Neuronal birth to cortical circuitry.Science. 2013 May 31;340(6136):1058-9. doi: 10.1126/science.1235778. Science. 2013. PMID: 23723226 No abstract available.
References
-
- Szentágothai J, Arbib MA. Conceptual models of neural organization. Neurosci Res Program Bull. 1974;12:305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
