

Prospective cognition in rats
- PMID: 23180886
- PMCID: PMC3501753
- DOI: 10.1016/j.lmot.2012.05.006
Prospective cognition in rats
Abstract
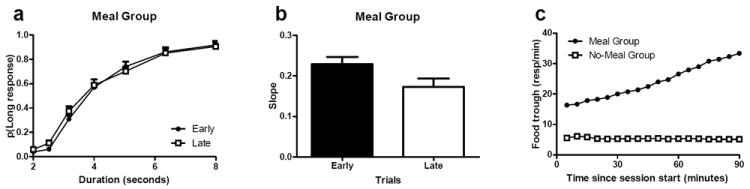
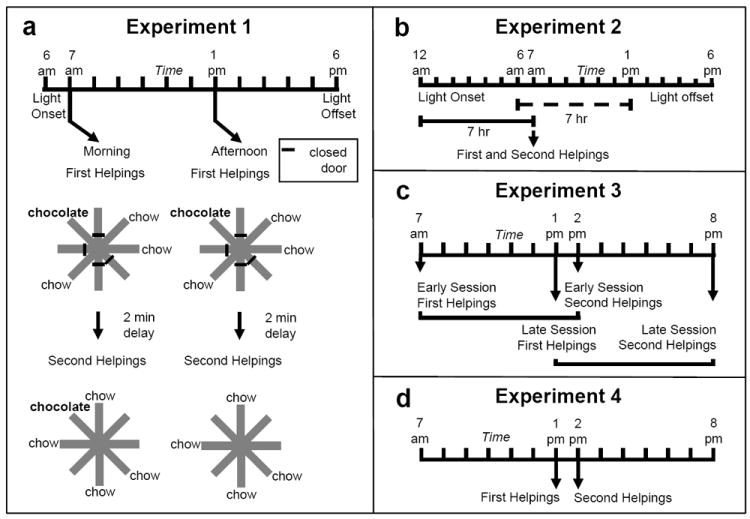
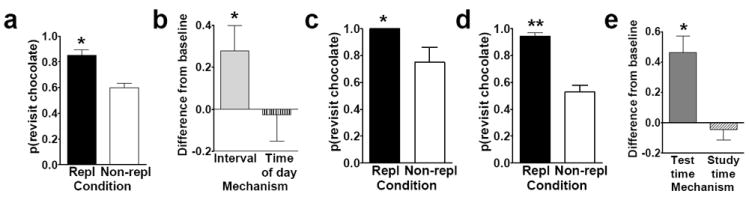
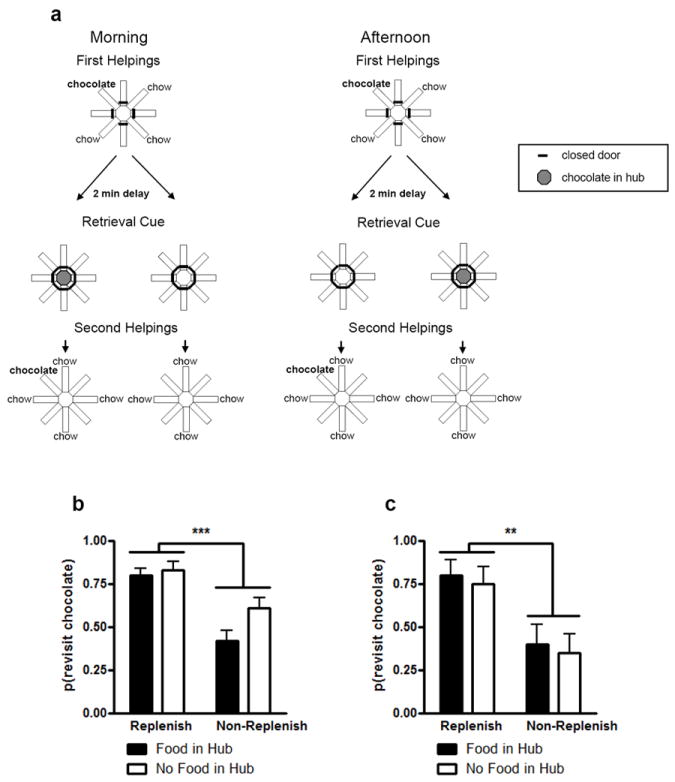
Efforts to develop animal models of memory are critical for understanding the neural substrate of memory. Memory is essential for daily life and enables information to be stored and retrieved after seconds to years. The ability to remember episodes from the past is thought to be related to the ability to plan for the future. Here we focus on a particular aspect of prospective cognition, namely the ability to remember to take action when a future scenario occurs. This review focuses on a recently developed method to evaluate prospective memory in the rat. Available evidence suggests that rats remember to take action in the future, but little is known about the temporal specificity of such memories or about the flexibility and limitations of prospective memories. Recent studies that suggest that rats remember a specific past episode are reviewed to underscore potential approaches that may be used to explore the range and limits of prospective cognition. The review highlights some directions to explore, including the temporal specificity of prospective cognition, the range of flexibility or creativity within prospective cognition, and the constraints imposed by multiple motivational systems.
Figures
References
-
- Babb SJ, Crystal JD. Discrimination of what, when, and where: Implications for episodic-like memory in rats. Learning & Motivation. 2005;36:177–189.
-
- Babb SJ, Crystal JD. Discrimination of what, when, and where is not based on time of day. Learning & Behavior. 2006a;34:124–130. - PubMed
-
- Babb SJ, Crystal JD. Episodic-like memory in the rat. Current Biology. 2006b;16:1317–1321. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous