

Regulation of primitive hematopoiesis by class I histone deacetylases
- PMID: 23184530
- PMCID: PMC3553261
- DOI: 10.1002/dvdy.23906
Regulation of primitive hematopoiesis by class I histone deacetylases
Abstract
Background: Histone deacetylases (HDACs) regulate multiple developmental processes and cellular functions. However, their roles in blood development have not been determined, and in Xenopus laevis a specific function for HDACs has yet to be identified. Here, we employed the class I selective HDAC inhibitor, valproic acid (VPA), to show that HDAC activity is required for primitive hematopoiesis.
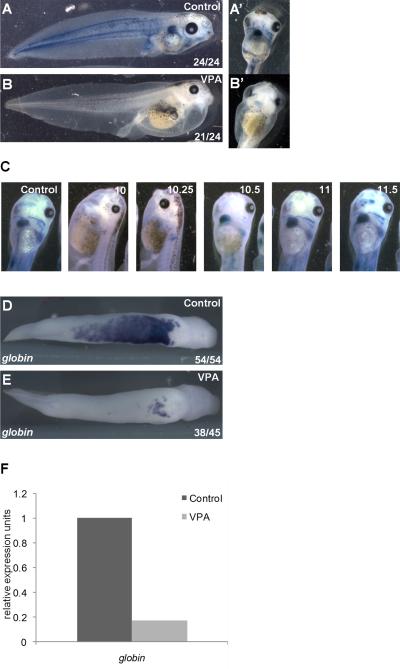
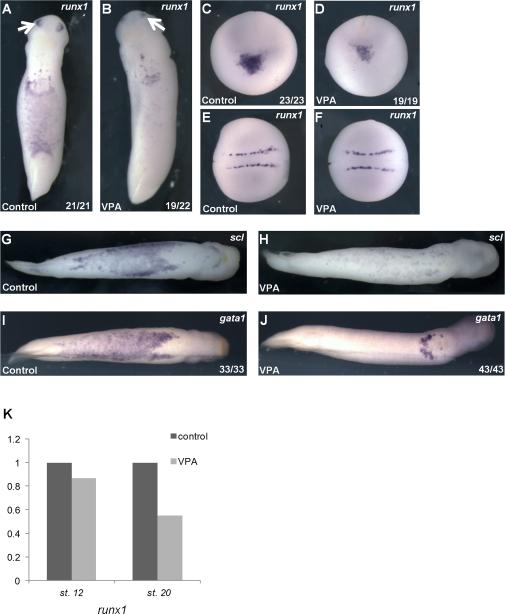
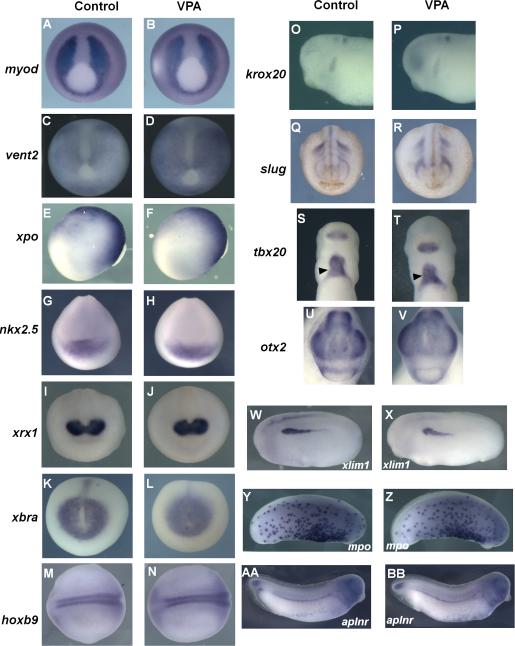
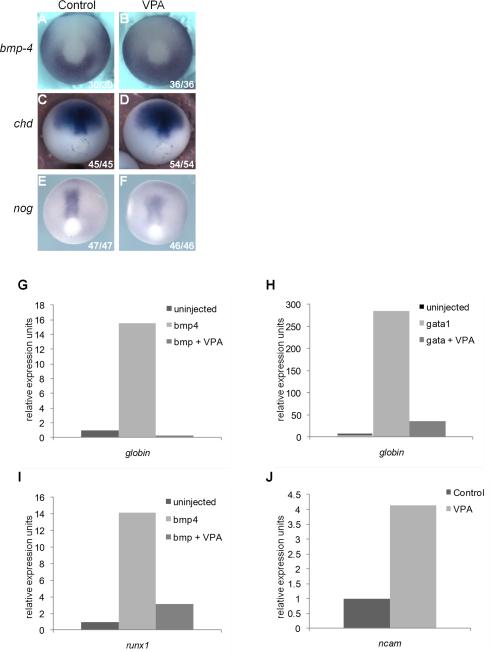
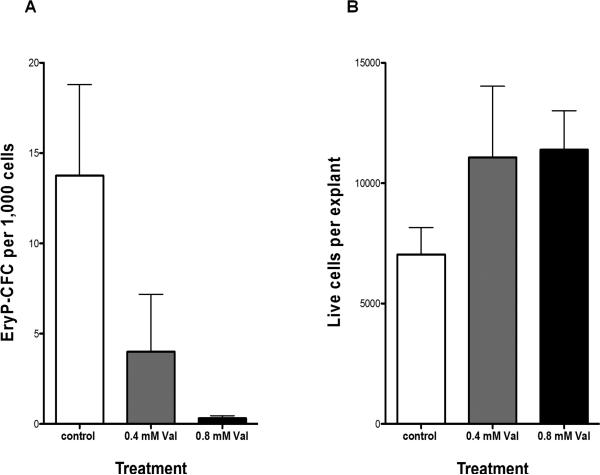
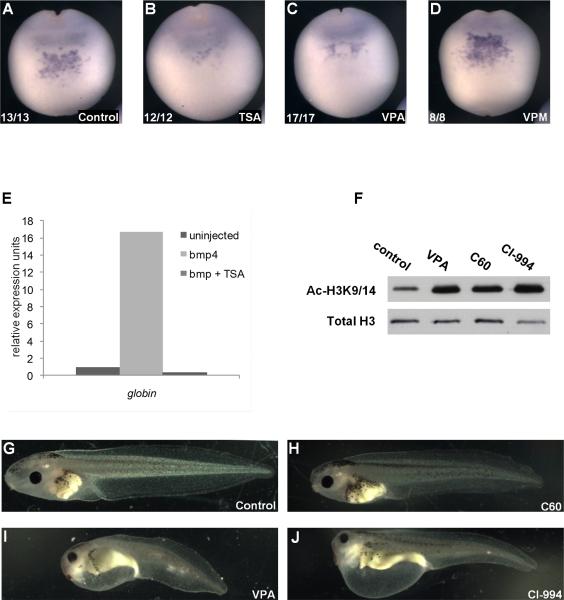
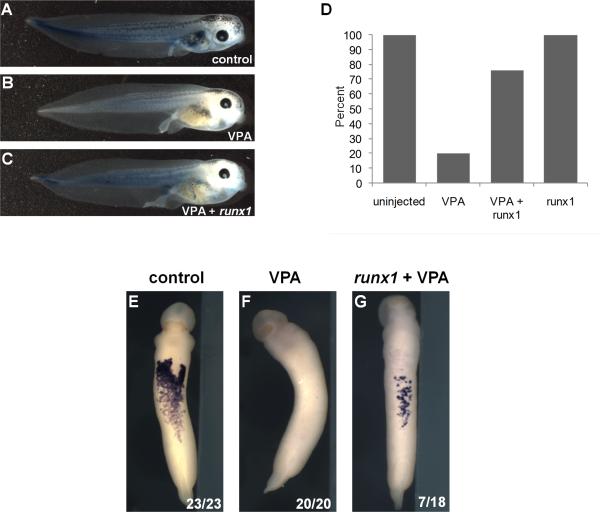
Results: VPA treatment during gastrulation resulted in a complete absence of red blood cells (RBCs) in Xenopus tadpoles, but did not affect development of other mesodermal tissues, including myeloid and endothelial lineages. These effects of VPA were mimicked by Trichostatin A (TSA), a well-established pan-HDAC inhibitor, but not by valpromide, which is structurally similar to VPA but does not inhibit HDACs. VPA also caused a marked, dose-dependent loss of primitive erythroid progenitors in mouse yolk sac explants at clinically relevant concentrations. In addition, VPA treatment inhibited erythropoietic development downstream of bmp4 and gata1 in Xenopus ectodermal explants.
Conclusions: These findings suggest an important role for class I HDACs in primitive hematopoiesis. Our work also demonstrates that specific developmental defects associated with exposure to VPA, a significant teratogen in humans, arise through inhibition of class I HDACs.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
References
-
- Boyes J, Byfield P, Nakatani Y, Ogryzko V. Regulation of activity of the transcription factor GATA-1 by acetylation. Nature. 1998;396:594–598. - PubMed
-
- Ciau-Uitz A, Liu F, Patient R. Genetic control of hematopoietic development in Xenopus and zebrafish. Int J Dev Biol. 54:1139–1149. - PubMed
-
- Ciau-Uitz A, Walmsley M, Patient R. Distinct origins of adult and embryonic blood in Xenopus. Cell. 2000;102:787–796. - PubMed
-
- Cunliffe VT. Histone deacetylase 1 is required to repress Notch target gene expression during zebrafish neurogenesis and to maintain the production of motoneurones in response to hedgehog signalling. Development. 2004;131:2983–2995. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
