

The C terminus of the histone chaperone Asf1 cross-links to histone H3 in yeast and promotes interaction with histones H3 and H4
- PMID: 23184661
- PMCID: PMC3666882
- DOI: 10.1128/MCB.01053-12
The C terminus of the histone chaperone Asf1 cross-links to histone H3 in yeast and promotes interaction with histones H3 and H4
Abstract
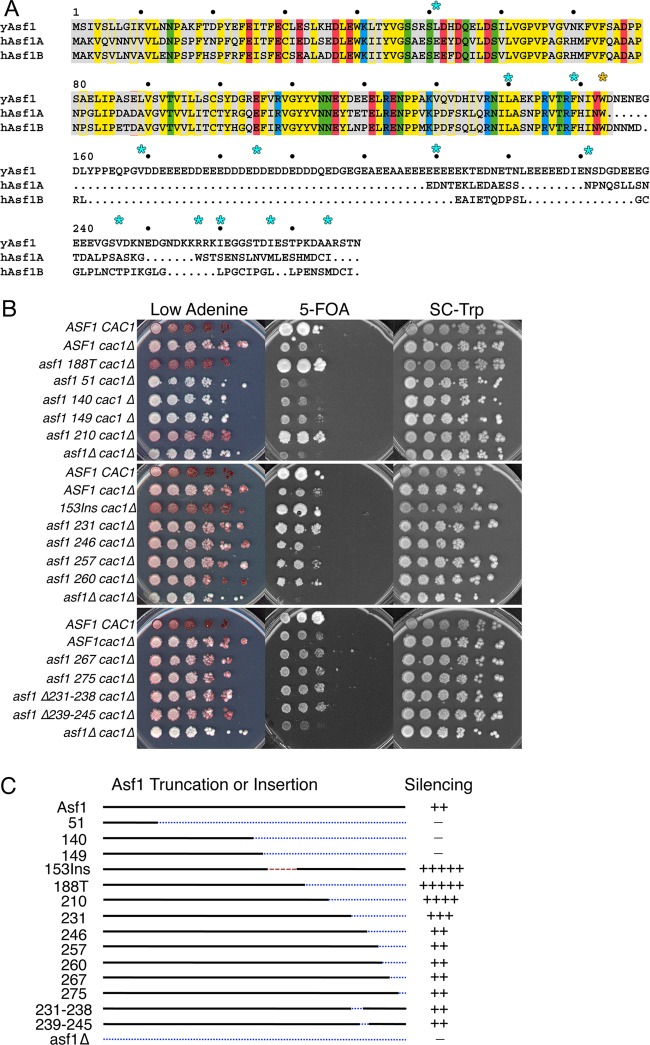
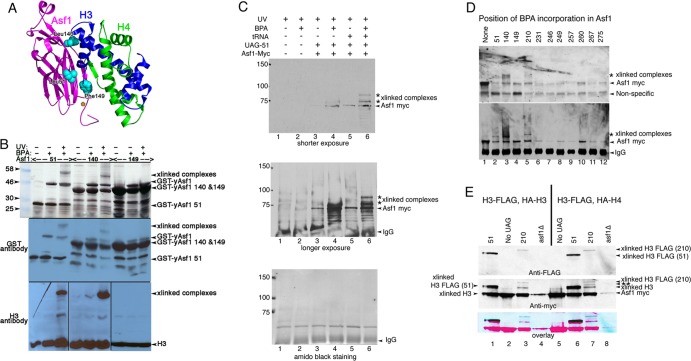
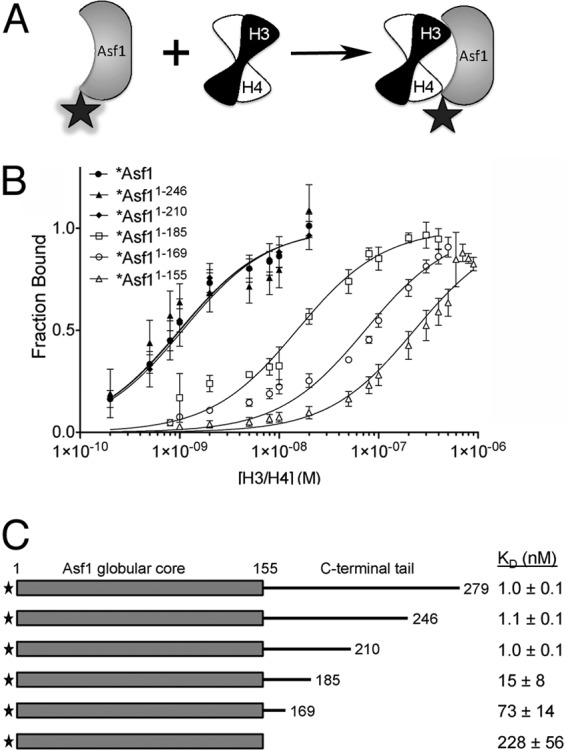
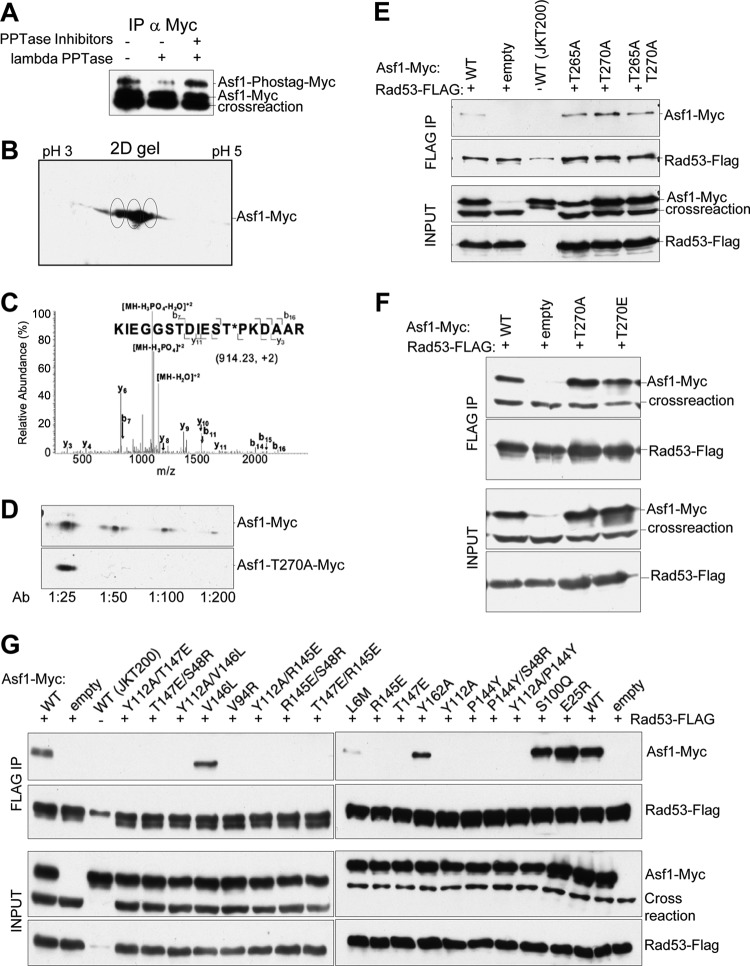
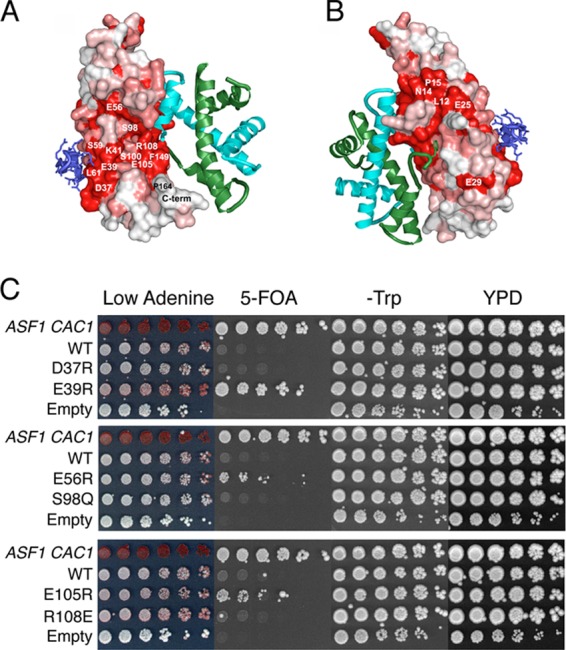
The central histone H3/H4 chaperone Asf1 comprises a highly conserved globular core and a divergent C-terminal tail. While the function and structure of the Asf1 core are well known, the function of the tail is less well understood. Here, we have explored the role of the yeast (yAsf1) and human (hAsf1a and hAsf1b) Asf1 tails in Saccharomyces cerevisiae. We show, using a photoreactive, unnatural amino acid, that Asf1 tail residue 210 cross-links to histone H3 in vivo and, further, that loss of C-terminal tail residues 211 to 279 weakens yAsf1-histone binding affinity in vitro nearly 200-fold. Via several yAsf1 C-terminal truncations and yeast-human chimeric proteins, we found that truncations at residue 210 increase transcriptional silencing and that the hAsf1a tail partially substitutes for full-length yAsf1 with respect to silencing but that full-length hAsf1b is a better overall substitute for full-length yAsf1. In addition, we show that the C-terminal tail of Asf1 is phosphorylated at T270 in yeast. Loss of this phosphorylation site does not prevent coimmunoprecipitation of yAsf1 and Rad53 from yeast extracts, whereas amino acid residue substitutions at the Asf1-histone H3/H4 interface do. Finally, we show that residue substitutions in yAsf1 near the CAF-1/HIRA interface also influence yAsf1's function in silencing.
Figures

Similar articles
-
Surprising complexity of the Asf1 histone chaperone-Rad53 kinase interaction.Proc Natl Acad Sci U S A. 2012 Feb 21;109(8):2866-71. doi: 10.1073/pnas.1106023109. Epub 2012 Feb 9. Proc Natl Acad Sci U S A. 2012. PMID: 22323608 Free PMC article.
-
Structural basis for the histone chaperone activity of Asf1.Cell. 2006 Nov 3;127(3):495-508. doi: 10.1016/j.cell.2006.08.047. Cell. 2006. PMID: 17081973 Free PMC article.
-
Dominant mutants of the Saccharomyces cerevisiae ASF1 histone chaperone bypass the need for CAF-1 in transcriptional silencing by altering histone and Sir protein recruitment.Genetics. 2006 Jun;173(2):599-610. doi: 10.1534/genetics.105.054783. Epub 2006 Apr 2. Genetics. 2006. PMID: 16582440 Free PMC article.
-
The histone chaperone Asf1 at the crossroads of chromatin and DNA checkpoint pathways.Chromosoma. 2007 Apr;116(2):79-93. doi: 10.1007/s00412-006-0087-z. Epub 2006 Dec 19. Chromosoma. 2007. PMID: 17180700 Review.
-
Understanding histone acetyltransferase Rtt109 structure and function: how many chaperones does it take?Curr Opin Struct Biol. 2011 Dec;21(6):728-34. doi: 10.1016/j.sbi.2011.09.005. Epub 2011 Oct 23. Curr Opin Struct Biol. 2011. PMID: 22023828 Free PMC article. Review.
Cited by
-
Mild dyserythropoiesis and β-like globin gene expression imbalance due to the loss of histone chaperone ASF1B.Hum Genomics. 2020 Oct 16;14(1):39. doi: 10.1186/s40246-020-00283-3. Hum Genomics. 2020. PMID: 33066815 Free PMC article.
-
Tousled-like kinases phosphorylate Asf1 to promote histone supply during DNA replication.Nat Commun. 2014 Mar 6;5:3394. doi: 10.1038/ncomms4394. Nat Commun. 2014. PMID: 24598821 Free PMC article.
-
The replisome guides nucleosome assembly during DNA replication.Cell Biosci. 2020 Mar 12;10:37. doi: 10.1186/s13578-020-00398-z. eCollection 2020. Cell Biosci. 2020. PMID: 32190287 Free PMC article. Review.
-
Unveiling novel interactions of histone chaperone Asf1 linked to TREX-2 factors Sus1 and Thp1.Nucleus. 2014 May-Jun;5(3):247-59. doi: 10.4161/nucl.29155. Epub 2014 May 13. Nucleus. 2014. PMID: 24824343 Free PMC article.
-
Fly Fishing for Histones: Catch and Release by Histone Chaperone Intrinsically Disordered Regions and Acidic Stretches.J Mol Biol. 2017 Aug 4;429(16):2401-2426. doi: 10.1016/j.jmb.2017.06.005. Epub 2017 Jun 10. J Mol Biol. 2017. PMID: 28610839 Free PMC article. Review.
References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. 1997. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389: 251– 260 - PubMed
-
- Le S, Davis C, Konopka JB, Sternglanz R. 1997. Two new S-phase-specific genes from Saccharomyces cerevisiae. Yeast 13: 1029– 1042 - PubMed
-
- Tyler JK, Adams CR, Chen SR, Kobayashi R, Kamakaka RT, Kadonaga JT. 1999. The RCAF complex mediates chromatin assembly during DNA replication and repair. Nature 402: 555– 560 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous