

Identification of a His-Asp-Cys catalytic triad essential for function of the Rho inactivation domain (RID) of Vibrio cholerae MARTX toxin
- PMID: 23184949
- PMCID: PMC3543022
- DOI: 10.1074/jbc.M112.396309
Identification of a His-Asp-Cys catalytic triad essential for function of the Rho inactivation domain (RID) of Vibrio cholerae MARTX toxin
Abstract
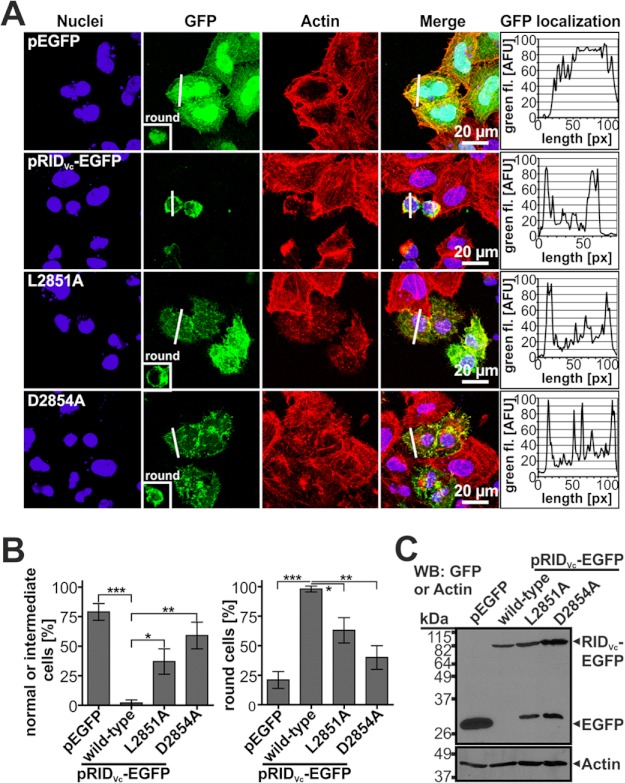
Vibrio cholerae is the causative agent of the severe diarrheal disease cholera. For V. cholerae to colonize the intestinal epithelium, accessory toxins such as the multifunctional autoprocessing repeats-in-toxin (MARTX(Vc)) toxin are required. MARTX toxins are composite toxins comprised of arrayed effector domains that carry out distinct functions inside the host cell. Among the three effector domains of MARTX(Vc) is the Rho inactivation domain (RID(Vc)) known to cause cell rounding through inactivation of small RhoGTPases. Using alanine scanning mutagenesis in the activity subdomain of RID(Vc), four residues, His-2782, Leu-2851, Asp-2854, and Cys-3022, were identified as impacting RID(Vc) function in depolymerization of the actin cytoskeleton and inactivation of RhoA. Tyr-2807 and Tyr-3015 were identified as important potentially for forming the active structure for substrate contact but are not involved in catalysis or post translational modifications. Finally, V. cholerae strains modified to carry a catalytically inactive RID(Vc) show that the rate and efficiency of MARTX(Vc) actin cross-linking activity does not depend on a functional RID(Vc), demonstrating that these domains function independently in actin depolymerization. Overall, our results indicate a His-Asp-Cys catalytic triad is essential for function of the RID effector domain family shared by MARTX toxins produced by many Gram-negative bacteria.
Figures








References
-
- Satchell K. J. (2011) Structure and function of MARTX toxins and other large repetitive RTX proteins. Annu. Rev. Microbiol. 65, 71–90 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
