

Nonmedially assembled F-actin cables incorporate into the actomyosin ring in fission yeast
- PMID: 23185032
- PMCID: PMC3514790
- DOI: 10.1083/jcb.201209044
Nonmedially assembled F-actin cables incorporate into the actomyosin ring in fission yeast
Abstract
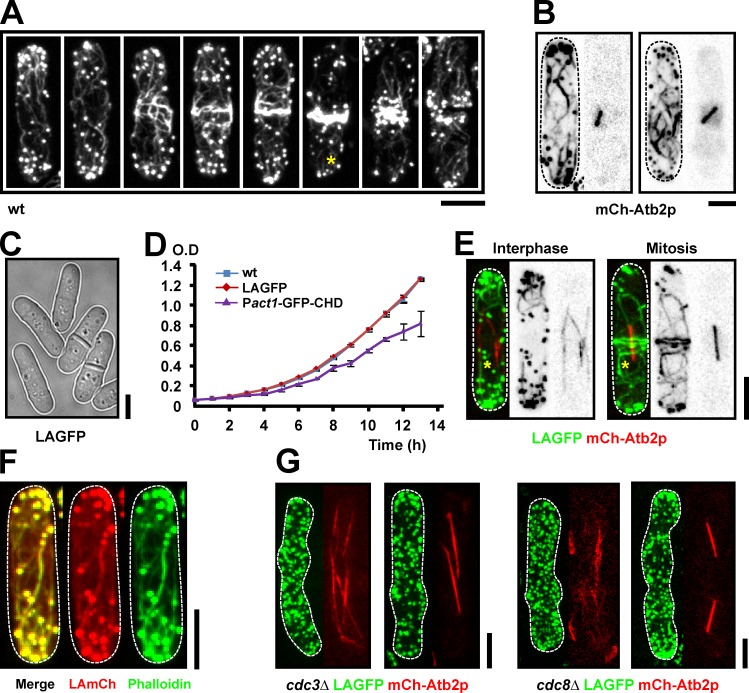
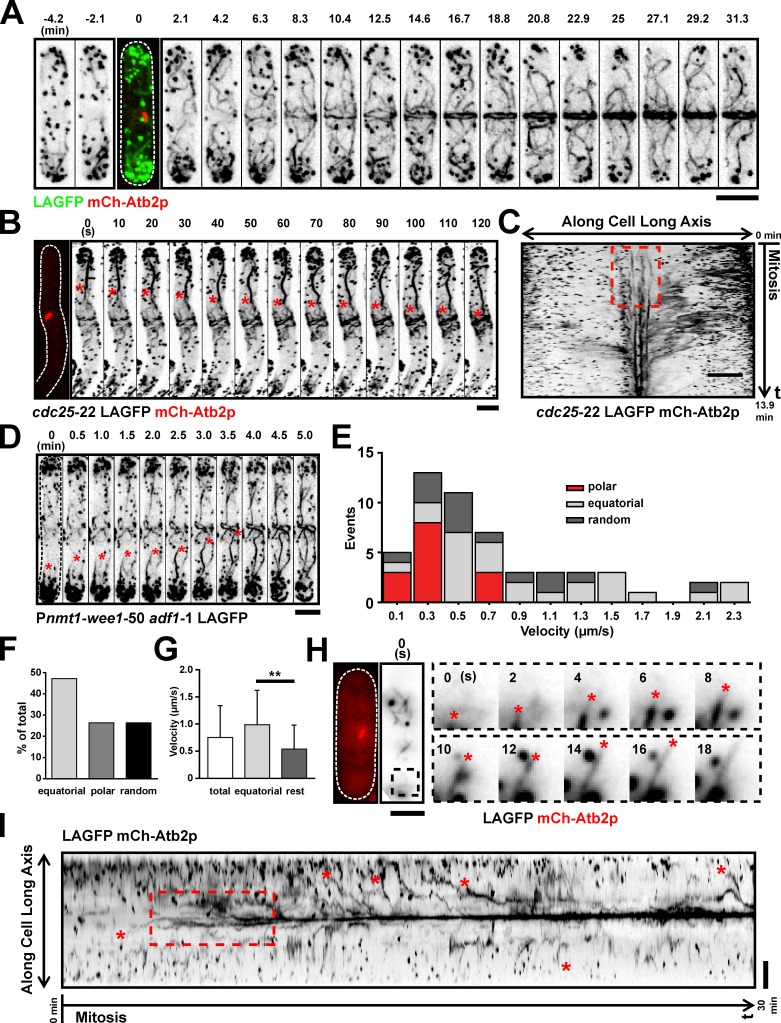
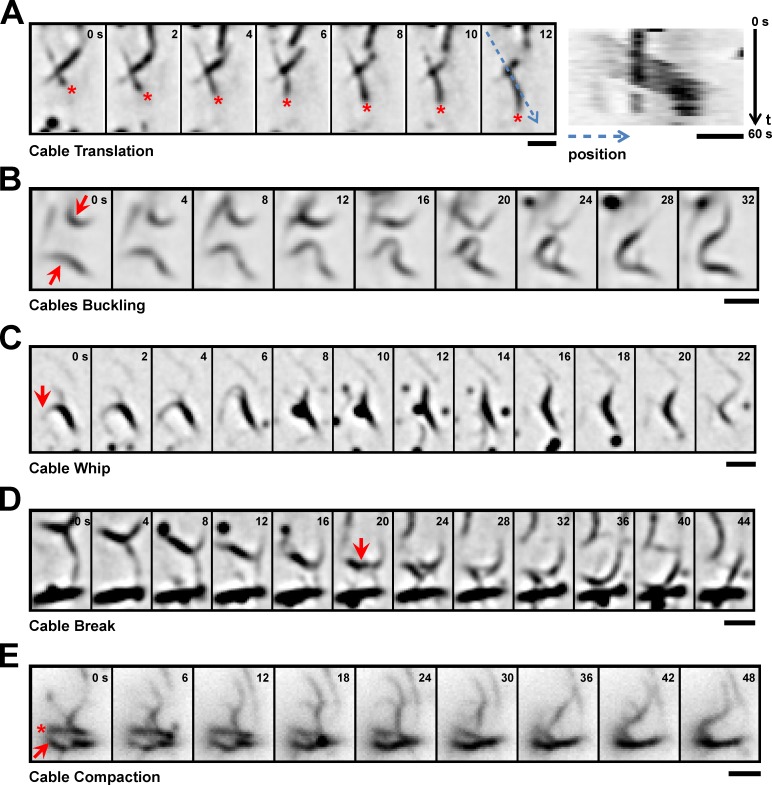
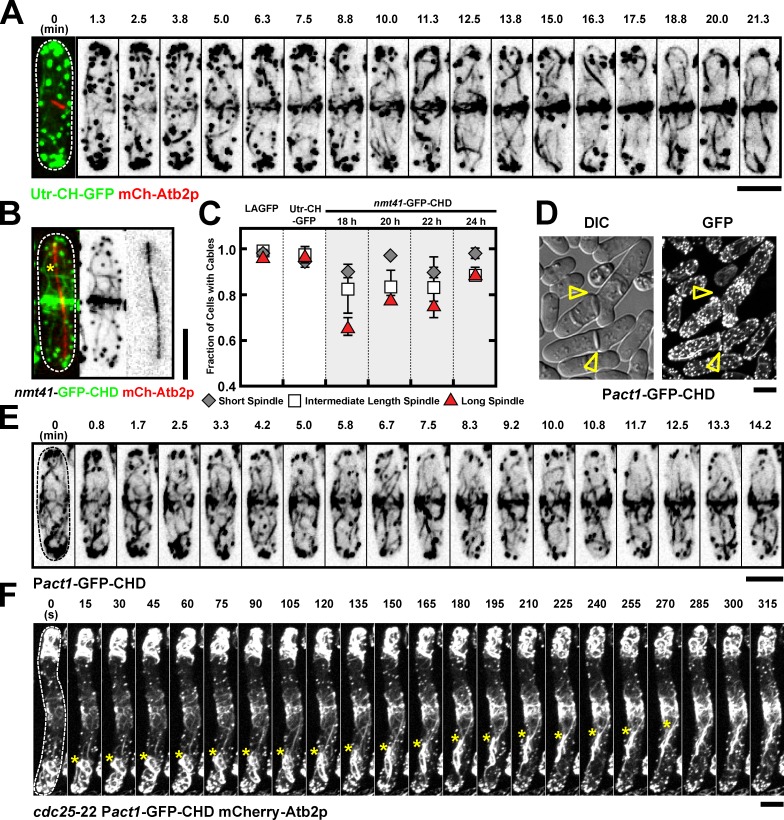
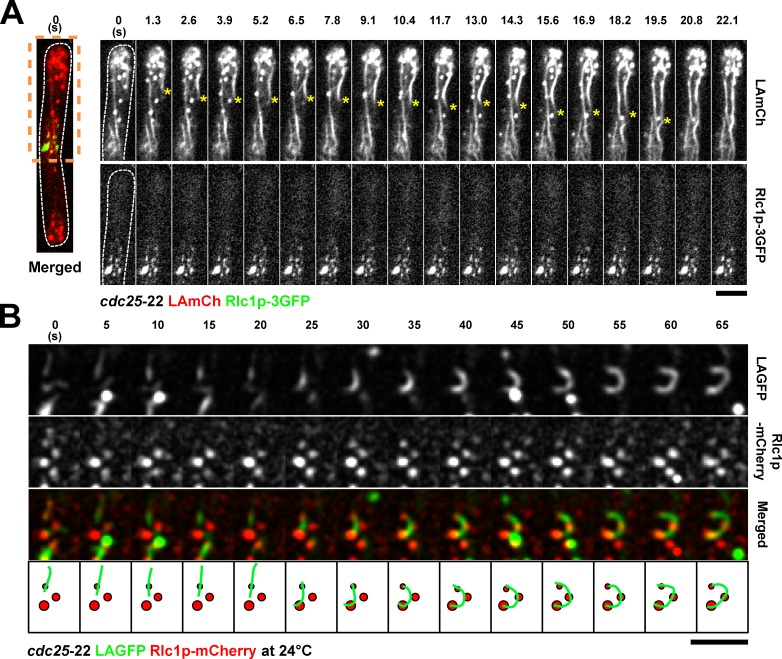
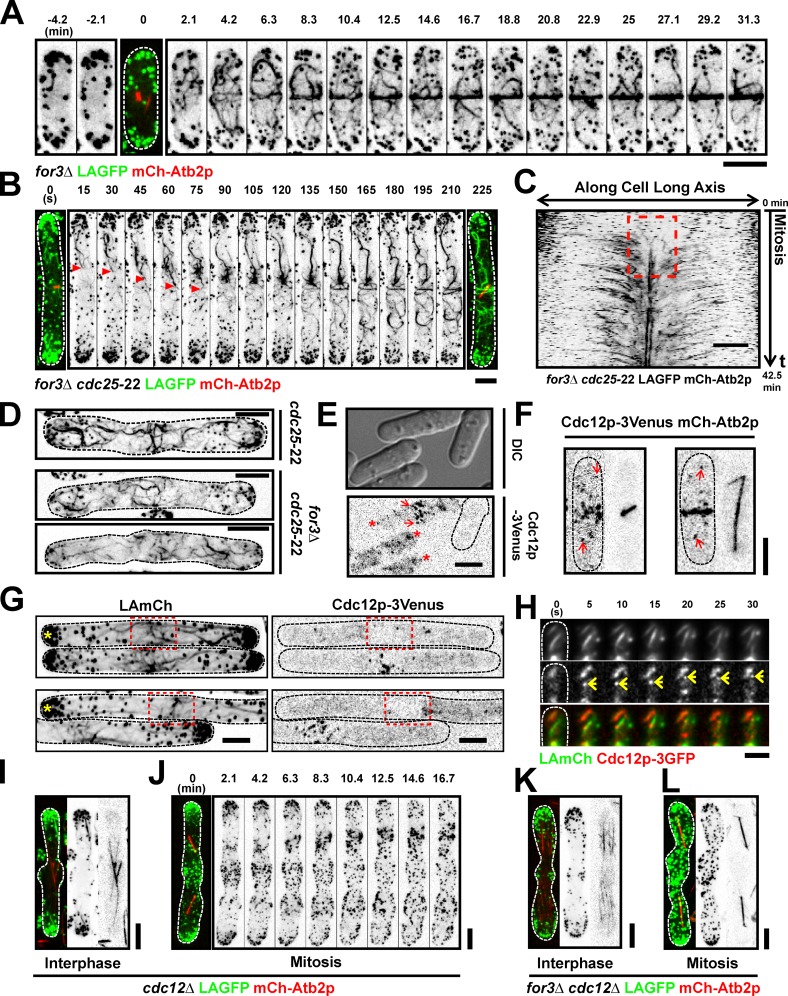
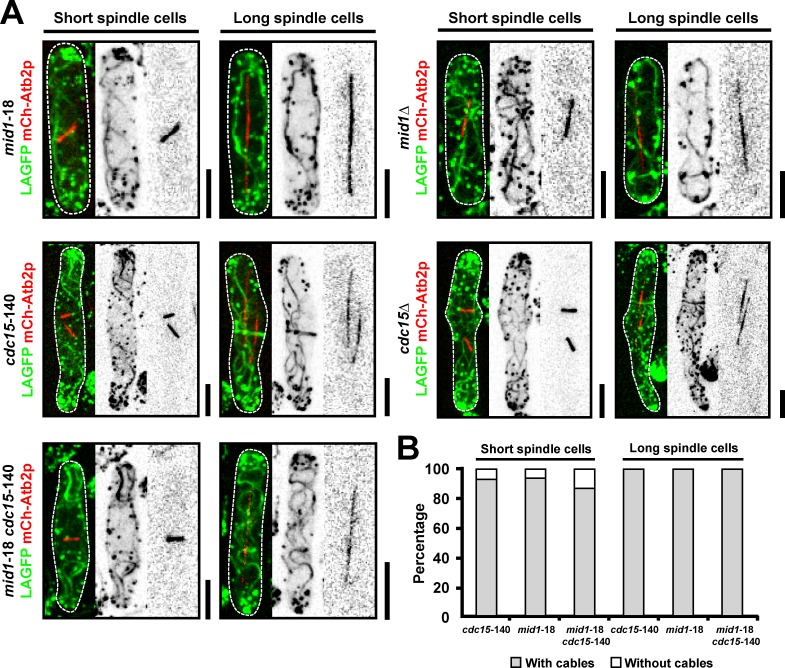
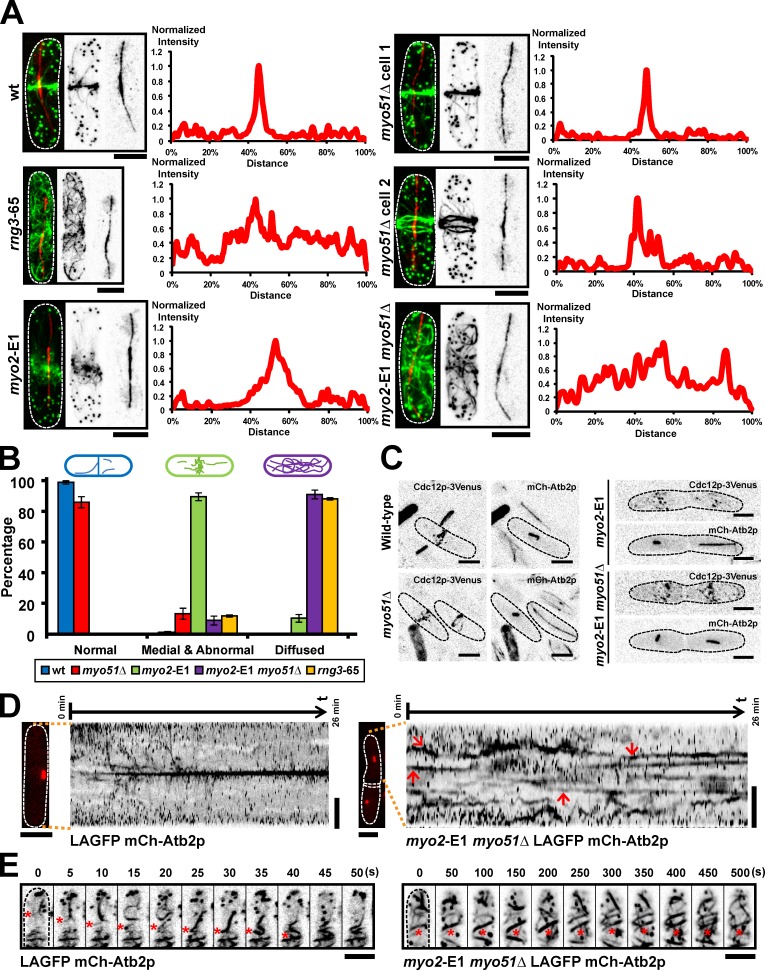
In many eukaryotes, cytokinesis requires the assembly and constriction of an actomyosin-based contractile ring. Despite the central role of this ring in cytokinesis, the mechanism of F-actin assembly and accumulation in the ring is not fully understood. In this paper, we investigate the mechanism of F-actin assembly during cytokinesis in Schizosaccharomyces pombe using lifeact as a probe to monitor actin dynamics. Previous work has shown that F-actin in the actomyosin ring is assembled de novo at the division site. Surprisingly, we find that a significant fraction of F-actin in the ring was recruited from formin-Cdc12p nucleated long actin cables that were generated at multiple nonmedial locations and incorporated into the ring by a combination of myosin II and myosin V activities. Our results, together with findings in animal cells, suggest that de novo F-actin assembly at the division site and directed transport of F-actin cables assembled elsewhere can contribute to ring assembly.
Figures
References
-
- Arai R., Mabuchi I. 2002. F-actin ring formation and the role of F-actin cables in the fission yeast Schizosaccharomyces pombe. J. Cell Sci. 115:887–898 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
