

H(2) enhances arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion
- PMID: 23185443
- PMCID: PMC3504229
- DOI: 10.1371/journal.pone.0049800
H(2) enhances arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion
Abstract
Background: The metabolism of hydrogen gas (H(2)) in bacteria and algae has been extensively studied for the interesting of developing H(2)-based fuel. Recently, H(2) is recognized as a therapeutic antioxidant and activates several signalling pathways in clinical trials. However, underlying physiological roles and mechanisms of H(2) in plants as well as its signalling cascade remain unknown.
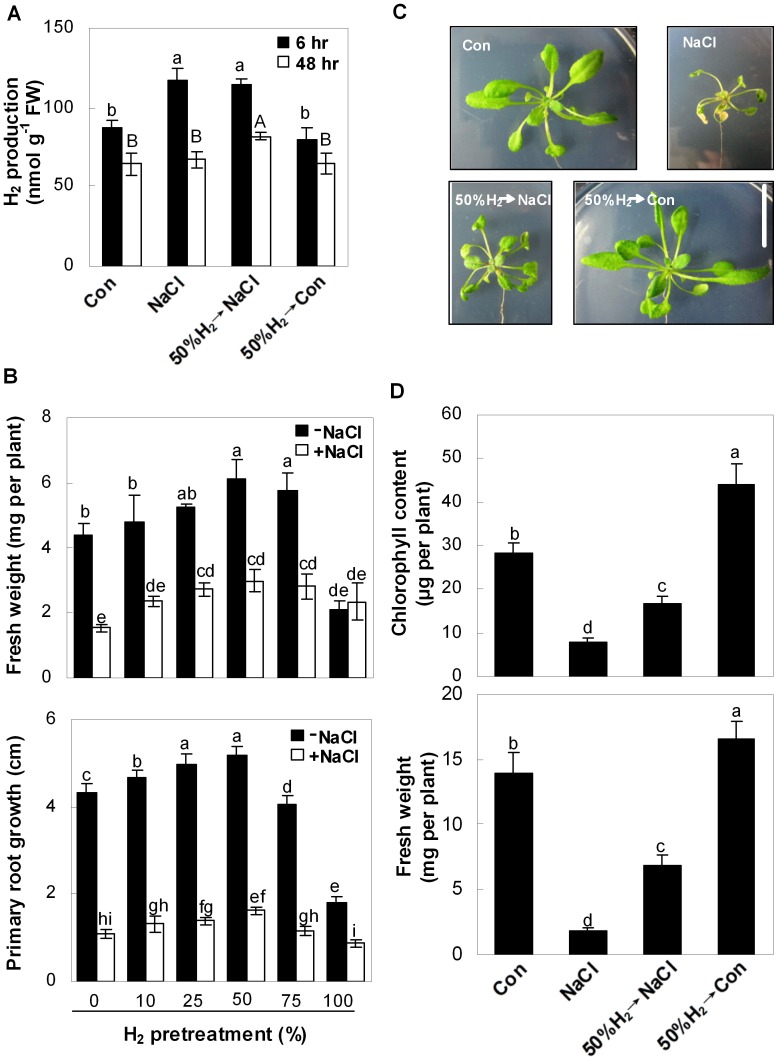
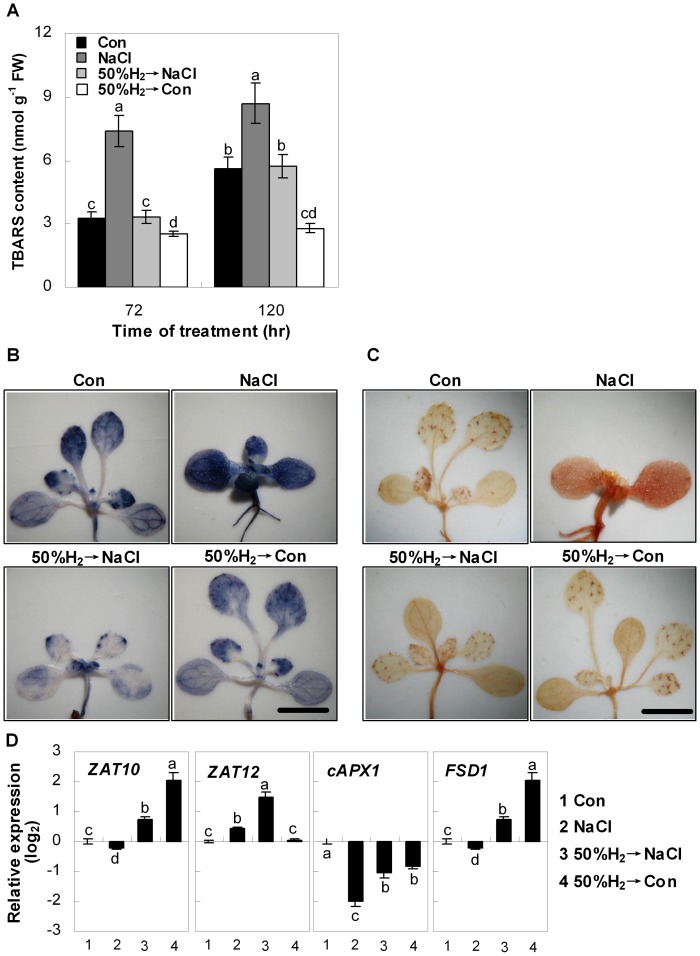
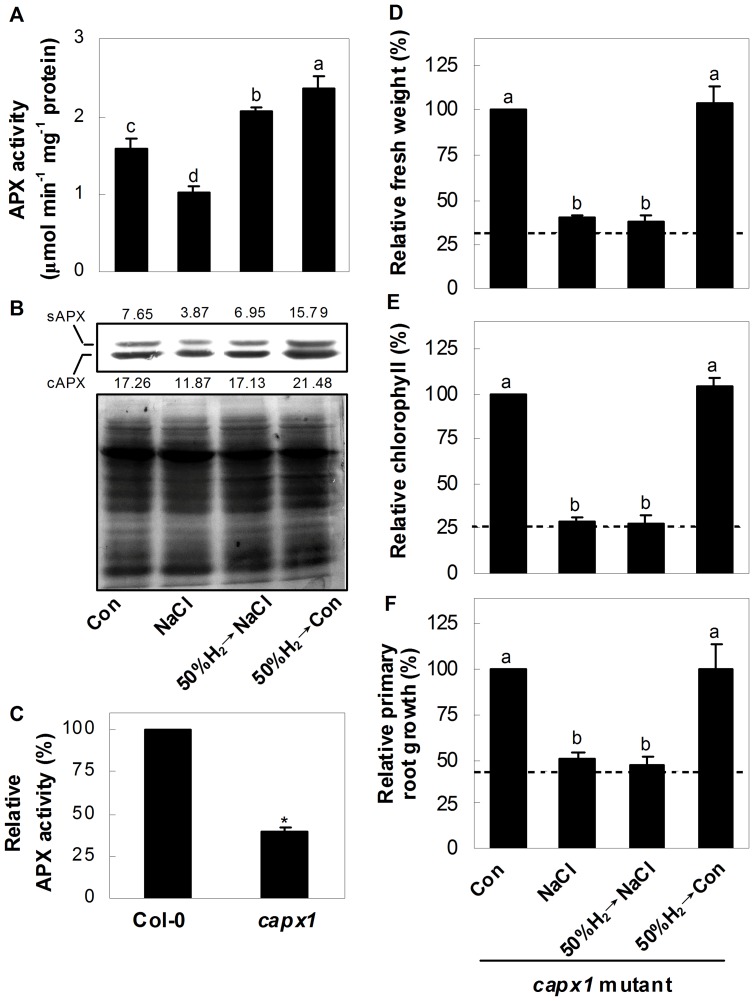
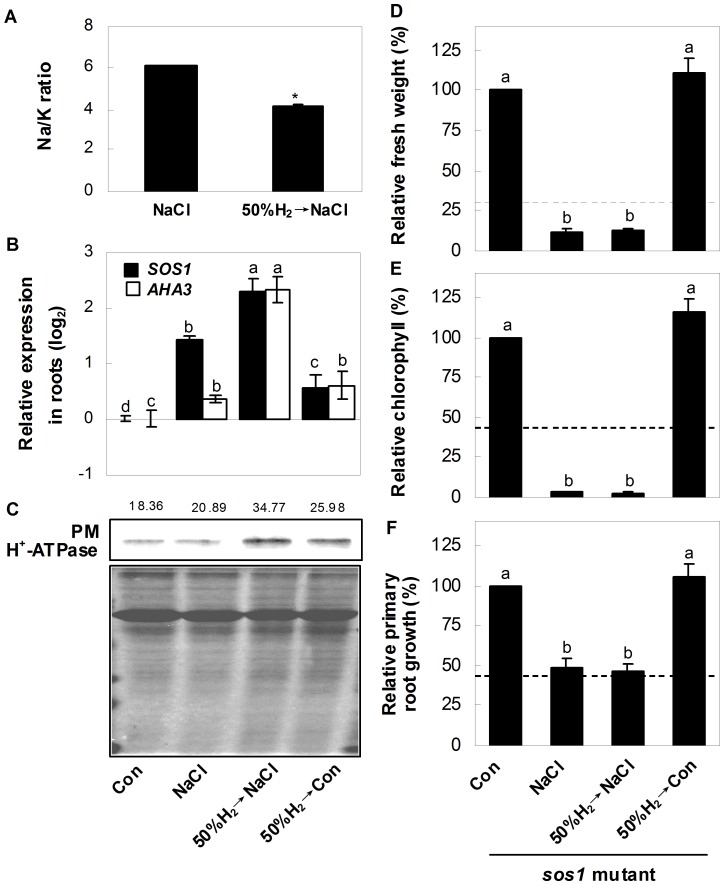
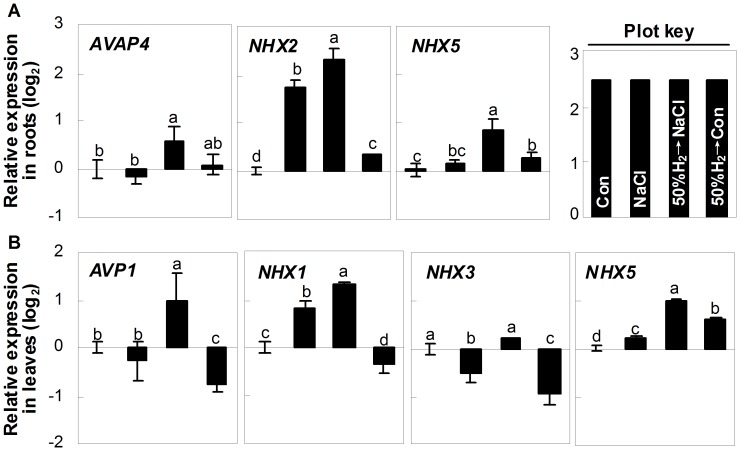
Methodology/principal findings: In this report, histochemical, molecular, immunological and genetic approaches were applied to characterize the participation of H(2) in enhancing Arabidopsis salt tolerance. An increase of endogenous H(2) release was observed 6 hr after exposure to 150 mM NaCl. Arabidopsis pretreated with 50% H(2)-saturated liquid medium, mimicking the induction of endogenous H(2) release when subsequently exposed to NaCl, effectively decreased salinity-induced growth inhibition. Further results showed that H(2) pretreatment modulated genes/proteins of zinc-finger transcription factor ZAT10/12 and related antioxidant defence enzymes, thus significantly counteracting the NaCl-induced reactive oxygen species (ROS) overproduction and lipid peroxidation. Additionally, H(2) pretreatment maintained ion homeostasis by regulating the antiporters and H(+) pump responsible for Na(+) exclusion (in particular) and compartmentation. Genetic evidence suggested that SOS1 and cAPX1 might be the target genes of H(2) signalling.
Conclusions: Overall, our findings indicate that H(2) acts as a novel and cytoprotective regulator in coupling ZAT10/12-mediated antioxidant defence and maintenance of ion homeostasis in the improvement of Arabidopsis salt tolerance.
Conflict of interest statement
Figures
Similar articles
-
The AtrbohF-dependent regulation of ROS signaling is required for melatonin-induced salinity tolerance in Arabidopsis.Free Radic Biol Med. 2017 Jul;108:465-477. doi: 10.1016/j.freeradbiomed.2017.04.009. Epub 2017 Apr 12. Free Radic Biol Med. 2017. PMID: 28412199
-
Molecular mechanisms of CAX3 involved in salt tolerance in Arabidopsis.Plant Physiol Biochem. 2025 Jun;223:109902. doi: 10.1016/j.plaphy.2025.109902. Epub 2025 Apr 10. Plant Physiol Biochem. 2025. PMID: 40215736
-
Hydrogen gas enhances Arabidopsis salt tolerance by modulating hydrogen peroxide-mediated redox and ion homeostasis.Plant Sci. 2025 Mar;352:112356. doi: 10.1016/j.plantsci.2024.112356. Epub 2024 Dec 7. Plant Sci. 2025. PMID: 39653253
-
A nuclear calcium-sensing pathway is critical for gene regulation and salt stress tolerance in Arabidopsis.PLoS Genet. 2013 Aug;9(8):e1003755. doi: 10.1371/journal.pgen.1003755. Epub 2013 Aug 29. PLoS Genet. 2013. PMID: 24009530 Free PMC article.
-
Plants pass the salt.Trends Plant Sci. 2003 May;8(5):200-1. doi: 10.1016/S1360-1385(03)00059-1. Trends Plant Sci. 2003. PMID: 12758034 Review.
Cited by
-
Genome-wide identification of the C2H2 zinc finger gene family and expression analysis under salt stress in sweetpotato.Front Plant Sci. 2023 Dec 13;14:1301848. doi: 10.3389/fpls.2023.1301848. eCollection 2023. Front Plant Sci. 2023. PMID: 38152142 Free PMC article.
-
Transcriptome Analysis of mRNA and miRNA in Somatic Embryos of Larix leptolepis Subjected to Hydrogen Treatment.Int J Mol Sci. 2016 Nov 22;17(11):1951. doi: 10.3390/ijms17111951. Int J Mol Sci. 2016. PMID: 27879674 Free PMC article.
-
Molecular hydrogen: a preventive and therapeutic medical gas for various diseases.Oncotarget. 2017 Sep 21;8(60):102653-102673. doi: 10.18632/oncotarget.21130. eCollection 2017 Nov 24. Oncotarget. 2017. PMID: 29254278 Free PMC article. Review.
-
Regulation of hydrogen rich water on strawberry seedlings and root endophytic bacteria under salt stress.Front Plant Sci. 2024 Nov 21;15:1497362. doi: 10.3389/fpls.2024.1497362. eCollection 2024. Front Plant Sci. 2024. PMID: 39640989 Free PMC article.
-
The molecular framework balancing growth and defense in response to plant elicitor peptide-induced signals in Arabidopsis.Plant Cell. 2024 Dec 23;37(1):koae327. doi: 10.1093/plcell/koae327. Plant Cell. 2024. PMID: 39700410 Free PMC article.
References
-
- Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6: 66–71. - PubMed
-
- Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59: 651–681. - PubMed
-
- Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7: 405–410. - PubMed
-
- Mittler R, Vanderauwera S, Gollery M, Breusegem FV (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9: 490–498. - PubMed
-
- Miller G, Suzuki N, Clftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ 33: 453–467. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
