

Stimulation of cortical myosin phosphorylation by p114RhoGEF drives cell migration and tumor cell invasion
- PMID: 23185572
- PMCID: PMC3501466
- DOI: 10.1371/journal.pone.0050188
Stimulation of cortical myosin phosphorylation by p114RhoGEF drives cell migration and tumor cell invasion
Abstract
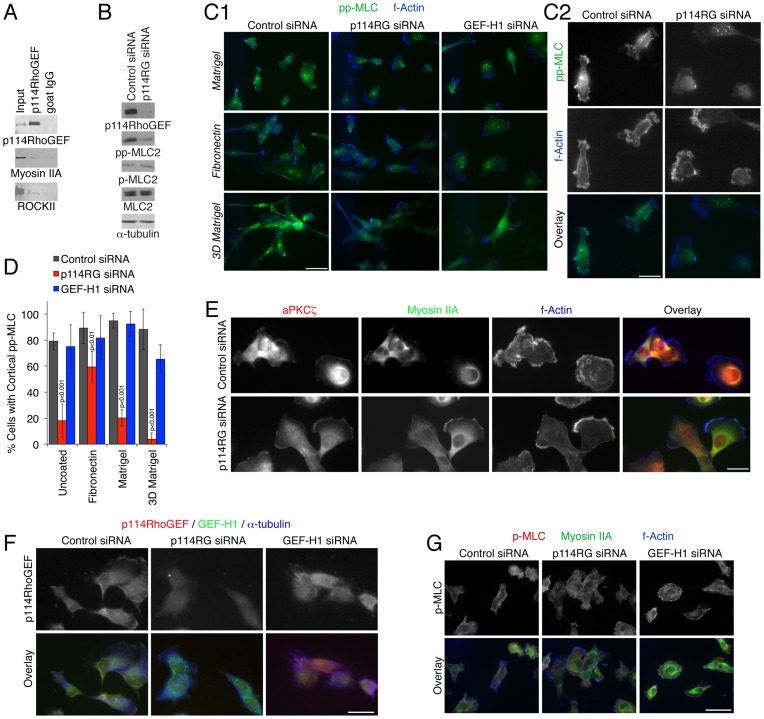
Actinomyosin activity is an important driver of cell locomotion and has been shown to promote collective cell migration of epithelial sheets as well as single cell migration and tumor cell invasion. However, the molecular mechanisms underlying activation of cortical myosin to stimulate single cell movement, and the relationship between the mechanisms that drive single cell locomotion and those that mediate collective cell migration of epithelial sheets are incompletely understood. Here, we demonstrate that p114RhoGEF, an activator of RhoA that associates with non-muscle myosin IIA, regulates collective cell migration of epithelial sheets and tumor cell invasion. Depletion of p114RhoGEF resulted in specific spatial inhibition of myosin activation at cell-cell contacts in migrating epithelial sheets and the cortex of migrating single cells, but only affected double and not single phosphorylation of myosin light chain. In agreement, overall elasticity and contractility of the cells, processes that rely on persistent and more constant forces, were not affected, suggesting that p114RhoGEF mediates process-specific myosin activation. Locomotion was p114RhoGEF-dependent on Matrigel, which favors more roundish cells and amoeboid-like actinomyosin-driven movement, but not on fibronectin, which stimulates flatter cells and lamellipodia-driven, mesenchymal-like migration. Accordingly, depletion of p114RhoGEF led to reduced RhoA, but increased Rac activity. Invasion of 3D matrices was p114RhoGEF-dependent under conditions that do not require metalloproteinase activity, supporting a role of p114RhoGEF in myosin-dependent, amoeboid-like locomotion. Our data demonstrate that p114RhoGEF drives cortical myosin activation by stimulating myosin light chain double phosphorylation and, thereby, collective cell migration of epithelial sheets and amoeboid-like motility of tumor cells.
Conflict of interest statement
Figures






Similar articles
-
Spatially restricted activation of RhoA signalling at epithelial junctions by p114RhoGEF drives junction formation and morphogenesis.Nat Cell Biol. 2011 Feb;13(2):159-66. doi: 10.1038/ncb2156. Epub 2011 Jan 23. Nat Cell Biol. 2011. PMID: 21258369 Free PMC article.
-
Vincristine enhances amoeboid-like motility via GEF-H1/RhoA/ROCK/Myosin light chain signaling in MKN45 cells.BMC Cancer. 2012 Oct 12;12:469. doi: 10.1186/1471-2407-12-469. BMC Cancer. 2012. PMID: 23057787 Free PMC article.
-
Intramolecular interactions between the Dbl homology (DH) domain and the carboxyl-terminal region of myosin II-interacting guanine nucleotide exchange factor (MyoGEF) act as an autoinhibitory mechanism for the regulation of MyoGEF functions.J Biol Chem. 2014 Dec 5;289(49):34033-48. doi: 10.1074/jbc.M114.607267. Epub 2014 Oct 21. J Biol Chem. 2014. PMID: 25336641 Free PMC article.
-
Ca2+ sensitivity of smooth muscle and nonmuscle myosin II: modulated by G proteins, kinases, and myosin phosphatase.Physiol Rev. 2003 Oct;83(4):1325-58. doi: 10.1152/physrev.00023.2003. Physiol Rev. 2003. PMID: 14506307 Review.
-
Amoeboid migration in health and disease: Immune responses versus cancer dissemination.Front Cell Dev Biol. 2023 Jan 5;10:1091801. doi: 10.3389/fcell.2022.1091801. eCollection 2022. Front Cell Dev Biol. 2023. PMID: 36699013 Free PMC article. Review.
Cited by
-
Activation of RhoA, but Not Rac1, Mediates Early Stages of S1P-Induced Endothelial Barrier Enhancement.PLoS One. 2016 May 17;11(5):e0155490. doi: 10.1371/journal.pone.0155490. eCollection 2016. PLoS One. 2016. PMID: 27187066 Free PMC article.
-
Genome-wide interrogation of extracellular vesicle biology using barcoded miRNAs.Elife. 2018 Dec 17;7:e41460. doi: 10.7554/eLife.41460. Elife. 2018. PMID: 30556811 Free PMC article.
-
Epithelial junctions and Rho family GTPases: the zonular signalosome.Small GTPases. 2014;5(4):1-15. doi: 10.4161/21541248.2014.973760. Small GTPases. 2014. PMID: 25483301 Free PMC article. Review.
-
GPCRs that Rhoar the Guanine nucleotide exchange factors.Small GTPases. 2022 Jan;13(1):84-99. doi: 10.1080/21541248.2021.1896963. Epub 2021 Apr 14. Small GTPases. 2022. PMID: 33849392 Free PMC article. Review.
-
Rho GTPases in collective cell migration.Small GTPases. 2014;5:e28997. doi: 10.4161/sgtp.28997. Epub 2014 May 9. Small GTPases. 2014. PMID: 25054920 Free PMC article. Review.
References
-
- Friedl P, Gilmour D (2009) Collective cell migration in morphogenesis, regeneration and cancer. Nat Rev Mol Cell Biol 10: 445–457. - PubMed
-
- Sanz-Moreno V, Marshall CJ (2010) The plasticity of cytoskeletal dynamics underlying neoplastic cell migration. Curr Opin Cell Biol 22: 690–696. - PubMed
-
- Lammermann T, Sixt M (2009) Mechanical modes of 'amoeboid' cell migration. Curr Opin Cell Biol 21: 636–644. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
