

Ethylene synthesis and regulated expression of recombinant protein in Synechocystis sp. PCC 6803
- PMID: 23185630
- PMCID: PMC3503970
- DOI: 10.1371/journal.pone.0050470
Ethylene synthesis and regulated expression of recombinant protein in Synechocystis sp. PCC 6803
Abstract
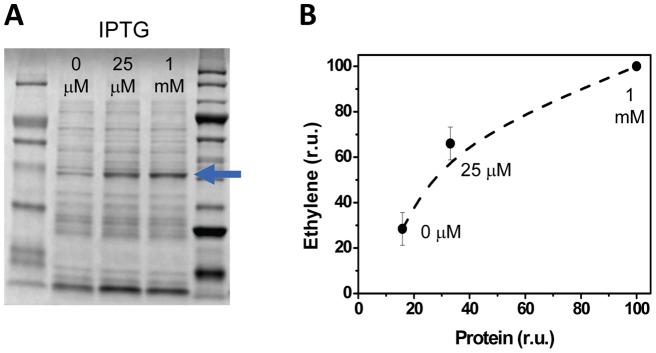
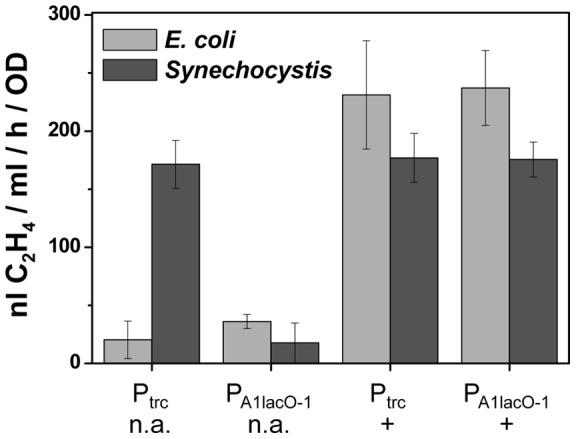
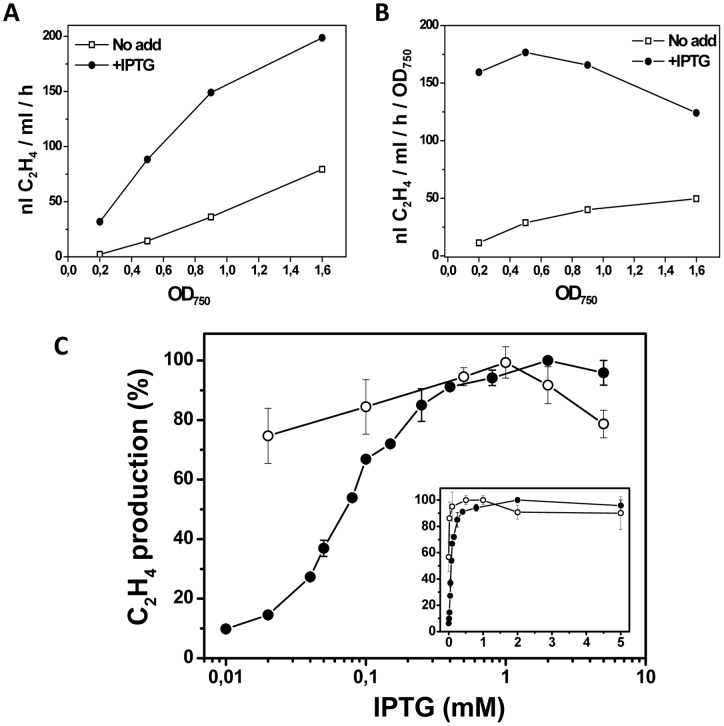
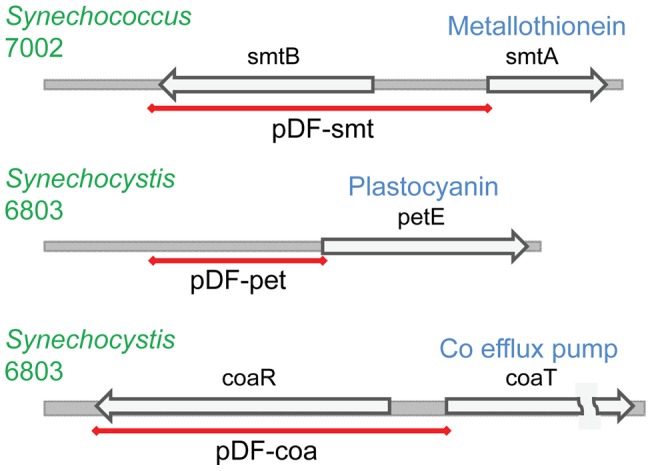
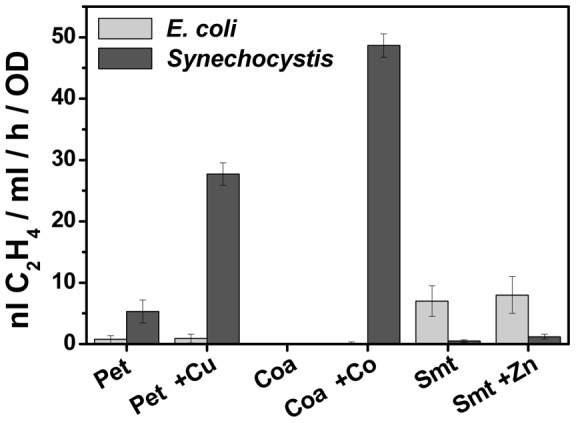
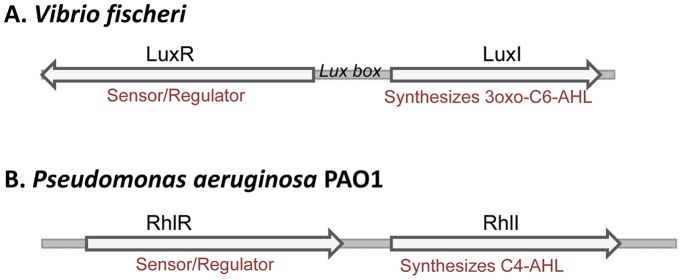
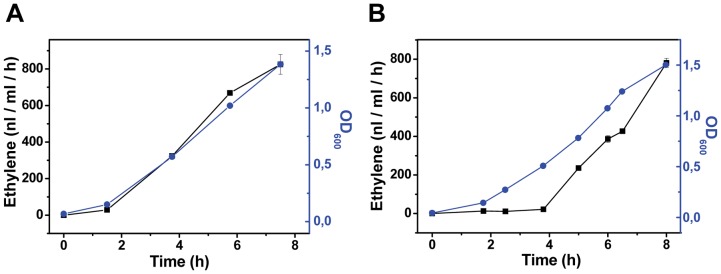
The ethylene-forming enzyme (EFE) from Pseudomonas syringae catalyzes the synthesis of ethylene which can be easily detected in the headspace of closed cultures. A synthetic codon-optimized gene encoding N-terminal His-tagged EFE (EFEh) was expressed in Synechocystis sp. PCC 6803 (Synechocystis) and Escherichia coli (E. coli) under the control of diverse promoters in a self-replicating broad host-range plasmid. Ethylene synthesis was stably maintained in both organisms in contrast to earlier work in Synechococcus elongatus PCC 7942. The rate of ethylene accumulation was used as a reporter for protein expression in order to assess promoter strength and inducibility with the different expression systems. Several metal-inducible cyanobacterial promoters did not function in E. coli but were well-regulated in cyanobacteria, albeit at a low level of expression. The E. coli promoter P(trc) resulted in constitutive expression in cyanobacteria regardless of whether IPTG was added or not. In contrast, a Lac promoter variant, P(A1lacO-1), induced EFE-expression in Synechocystis at a level of expression as high as the Trc promoter and allowed a fine level of IPTG-dependent regulation of protein-expression. The regulation was tight at low cell density and became more relaxed in more dense cultures. A synthetic quorum-sensing promoter system was also constructed and shown to function well in E. coli, however, only a very low level of EFE-activity was observed in Synechocystis, independent of cell density.
Conflict of interest statement
Figures


References
-
- Sakai M, Ogawa T, Matsuoka M, Fukuda H (1997) Photosynthetic Conversion of Carbon Dioxide to Ethylene by the Recombinant Cyanobacterium, Synechococcus sp. PCC 7942, Which Harbors a Gene for the Ethylene-Forming Enzyme of Pseudomonas syringae . Journal of Fermentation and Bioengineering 84: 434–443.
-
- Takahama K, Matsuoka M, Nagahama K, Ogawa T (2003) Construction and analysis of a recombinant cyanobacterium expressing a chromosomally inserted gene for an ethylene-forming enzyme at the psbAI locus. J Biosci Bioeng 95: 302–305. - PubMed
-
- Ungerer J, Tao L, Davis M, Ghirardi M, Maness P-C, et al. (2012) Sustained photosynthetic conversion of CO2 to ethylene in recombinant cyanobacterium Synechocystis 6803. Energy & Environmental Science.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
