

The 2.5 Å structure of the enterococcus conjugation protein TraM resembles VirB8 type IV secretion proteins
- PMID: 23188825
- PMCID: PMC3548508
- DOI: 10.1074/jbc.M112.428847
The 2.5 Å structure of the enterococcus conjugation protein TraM resembles VirB8 type IV secretion proteins
Abstract
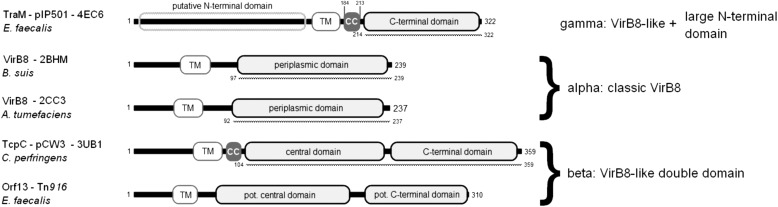
Conjugative plasmid transfer is the most important means of spreading antibiotic resistance and virulence genes among bacteria and therefore presents a serious threat to human health. The process requires direct cell-cell contact made possible by a multiprotein complex that spans cellular membranes and serves as a channel for macromolecular secretion. Thus far, well studied conjugative type IV secretion systems (T4SS) are of Gram-negative (G-) origin. Although many medically relevant pathogens (e.g., enterococci, staphylococci, and streptococci) are Gram-positive (G+), their conjugation systems have received little attention. This study provides structural information for the transfer protein TraM of the G+ broad host range Enterococcus conjugative plasmid pIP501. Immunolocalization demonstrated that the protein localizes to the cell wall. We then used opsonophagocytosis as a novel tool to verify that TraM was exposed on the cell surface. In these assays, antibodies generated to TraM recruited macrophages and enabled killing of pIP501 harboring Enteroccocus faecalis cells. The crystal structure of the C-terminal, surface-exposed domain of TraM was determined to 2.5 Å resolution. The structure, molecular dynamics, and cross-linking studies indicated that a TraM trimer acts as the biological unit. Despite the absence of sequence-based similarity, TraM unexpectedly displayed a fold similar to the T4SS VirB8 proteins from Agrobacterium tumefaciens and Brucella suis (G-) and to the transfer protein TcpC from Clostridium perfringens plasmid pCW3 (G+). Based on the alignments of secondary structure elements of VirB8-like proteins from mobile genetic elements and chromosomally encoded T4SS from G+ and G- bacteria, we propose a new classification scheme of VirB8-like proteins.
Figures




Similar articles
-
The conjugation protein TcpC from Clostridium perfringens is structurally related to the type IV secretion system protein VirB8 from Gram-negative bacteria.Mol Microbiol. 2012 Jan;83(2):275-88. doi: 10.1111/j.1365-2958.2011.07930.x. Epub 2011 Dec 11. Mol Microbiol. 2012. PMID: 22150951
-
The type IV secretion protein TraK from the Enterococcus conjugative plasmid pIP501 exhibits a novel fold.Acta Crystallogr D Biol Crystallogr. 2014 Apr;70(Pt 4):1124-35. doi: 10.1107/S1399004714001606. Epub 2014 Mar 21. Acta Crystallogr D Biol Crystallogr. 2014. PMID: 24699656
-
VirB8-like protein TraH is crucial for DNA transfer in Enterococcus faecalis.Sci Rep. 2016 Apr 22;6:24643. doi: 10.1038/srep24643. Sci Rep. 2016. PMID: 27103580 Free PMC article.
-
Conjugative type IV secretion systems in Gram-positive bacteria.Plasmid. 2013 Nov;70(3):289-302. doi: 10.1016/j.plasmid.2013.09.005. Epub 2013 Oct 12. Plasmid. 2013. PMID: 24129002 Free PMC article. Review.
-
VirB8: a conserved type IV secretion system assembly factor and drug target.Biochem Cell Biol. 2006 Dec;84(6):890-9. doi: 10.1139/o06-148. Biochem Cell Biol. 2006. PMID: 17215876 Review.
Cited by
-
Protein interactions within and between two F-type type IV secretion systems.Mol Microbiol. 2020 Nov;114(5):823-838. doi: 10.1111/mmi.14582. Epub 2020 Aug 12. Mol Microbiol. 2020. PMID: 32738086 Free PMC article.
-
Characterization of Cercospora nicotianae Hypothetical Proteins in Cercosporin Resistance.PLoS One. 2015 Oct 16;10(10):e0140676. doi: 10.1371/journal.pone.0140676. eCollection 2015. PLoS One. 2015. PMID: 26474162 Free PMC article.
-
Molecular and structural analysis of Legionella DotI gives insights into an inner membrane complex essential for type IV secretion.Sci Rep. 2015 Jun 3;5:10912. doi: 10.1038/srep10912. Sci Rep. 2015. PMID: 26039110 Free PMC article.
-
Elucidating assembly and function of VirB8 cell wall subunits refines the DNA translocation model in Gram-positive T4SSs.Sci Adv. 2025 Jan 24;11(4):eadq5975. doi: 10.1126/sciadv.adq5975. Epub 2025 Jan 22. Sci Adv. 2025. PMID: 39841841 Free PMC article.
-
The Rickettsia type IV secretion system: unrealized complexity mired by gene family expansion.Pathog Dis. 2016 Aug;74(6):ftw058. doi: 10.1093/femspd/ftw058. Epub 2016 Jun 14. Pathog Dis. 2016. PMID: 27307105 Free PMC article.
References
-
- Llosa M., Gomis-Rüth F. X., Coll M., de la Cruz F. (2002) Bacterial conjugation. A two-step mechanism for DNA transport. Mol. Microbiol. 45, 1–8 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
