

Review
doi: 10.1038/ncb2635.
Role of membrane traffic in the generation of epithelial cell asymmetry
Affiliations
- PMID: 23196841
- PMCID: PMC3771702
- DOI: 10.1038/ncb2635
Item in Clipboard
Review
Role of membrane traffic in the generation of epithelial cell asymmetry
Nat Cell Biol.
2012 Dec.
Abstract
Epithelial cells have an apical-basolateral axis of polarity, which is required for epithelial functions including barrier formation, vectorial ion transport and sensory perception. Here we review what is known about the sorting signals, machineries and pathways that maintain this asymmetry, and how polarity proteins interface with membrane-trafficking pathways to generate membrane domains de novo. It is becoming apparent that membrane traffic does not simply reinforce polarity, but is critical for the generation of cortical epithelial cell asymmetry.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Membrane-trafficking pathways in polarized epithelial cells. Proteins, lipids and fluid are internalized from the apical (step 1a) or basolateral (1b) surfaces, and delivered to apical early endosomes (AEEs) or basolateral early endosomes (BEEs). Cargoes can recycle to the cell surface (2a/b), be delivered to multivesicular bodies (MVBs) (3a/b), from which they will be delivered to lysosomes (not shown), or they can be delivered to the common recycling endosome (CRE; 4a/b). There may be a direct pathway from the AEE to the apical recycling endosome (ARE; 4c). CRE cargoes are shuttled to the basolateral cell surface (5), or delivered to ARE (6), from where they gain access to the apical cell surface (7a). Apical-to-basolateral transcytosis may involve cargo transit from the ARE to CRE (7b). On synthesis in the endoplasmic reticulum (not shown), proteins are delivered by way of the Golgi stacks to the trans-Golgi network (TGN), to take either direct routes to the apical (8) or basolateral (10) surfaces, or indirect routes through the ARE (9) or CRE (11). Endocytic pathways are shown with black arrows; biosynthetic ones with red arrows. The inset shows a magnified view of the apical junctional complex (JC), which is comprised of the zonula occludens (ZO), the zonula adherens (ZA) and the macula adherens (MA).

Use of two-dimensional versus three-dimensional models to study epithelial polarity. Epithelial cells can be grown under a number of conditions including on plastic dishes or coverslips (a), or on permeable filter supports such as Transwells (b). The latter, two-dimensional culture system is popular because it allows the epithelial cells to feed from their basolateral surfaces, they establish polarity within days, and the arrangement allows for access to both the apical and basolateral surfaces of the culture. An alternative to traditional two-dimensional culture involves growing epithelial tissues, cells or progenitors in a three-dimensional environment comprising extracellular matrix proteins, which allows for some aspects of tissue morphogenesis, such as lumen formation, to be recapitulated (c). For example, Lgr5-positive intestinal stem cells grown in three-dimensional cultures form ‘mini-guts’ lined by intestinal epithelial cells organized in the expected manner. Furthermore, when fragments of mammary glands, kidney and other organs are grown in such matrices they can recapitulate developmental processes such as branching morphogenesis, a phenotype not observed in two-dimensional cultures,. Moreover, when MDCK, MCF-10A or a number of other epithelial cell lines are cultured in a three-dimensional matrix, they form spheroid structures, variably referred to as cysts or acini, depending on the epithelial lineage of origin–. When mature, these spheroids comprise a single layer of epithelial cells with their apical surfaces lining an inner lumen, and their basolateral surface in contact with the extracellular matrix. When treated with morphogens some spheroids are able to undergo tubulogenetic programs, another process that cannot be modelled in two dimensions,.
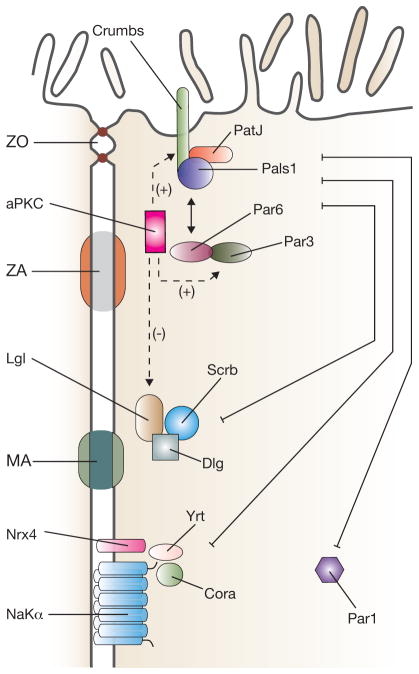
Polarity complexes in polarized vertebrate epithelial cells. The apical-most Crumbs complex includes the transmembrane protein crumbs and the cytoplasmic proteins Pals1 and PatJ. It interacts with the Par complex (aPKC, Par3 and Par6). aPKC phosphorylates crumbs and Par3, promoting apical identity. In contrast, aPKC-dependent phosphorylation of Lgl promotes its degradation, suppressing basolateral identity. Basolateral polarity is promoted by the Scribble complex (Scribble (Scrb), Discs large (Dlg) and Lethal giant larvae (Lgl)), Par1 and the Yurt (Yrt)–coracle (Cora) complex (Yrt, Cora, neurexin IV (Nrx4) and Na+/K+-ATPase α-subunit (NaKα)). The basolateral and apical polarity complexes act antagonistically to promote apical and basolateral membrane asymmetry. Components of the junctional complex are described in the legend to Fig. 1.

Early steps in lumen formation during MDCK cyst development. Before lumen formation, individual MDCK cells undergo mitosis. (a) At the two-cell stage polarization begins: E-cadherin, occludin and the exocyst subunit Sec10 are at the cell–cell adhesion, whereas Par3 and the exocyst subunit Sec8 are enriched at the periphery. The apical protein podocalyxin is found at the extracellular-matrix-abutting surface at this time. (b) During formation of the apical membrane initiation site (AMIS), part of the cell–cell contact is converted into an apical (luminal) surface. AMIS formation requires a series of trafficking events that include: endocytosis of podocalyxin and the crumbs complex from the periphery, and Rab11a-dependent transport to, and exocytosis at, the AMIS. Rab11a recruits a Fip5–Snx18 complex to promote vesicle formation from the apical recycling endosome (ARE). Rab11a also activates Rab8a/b by the GTP exchange factor (GEF) Rabin8 (opposed by the Rab8 GTPase activating protein (GAP) Tbc1d30). Rab8a and 11a recruit Myo5b and the Sec15a exocyst subunit to transport vesicles to the AMIS, and interact with Sec10 at the AMIS to promote vesicle docking. A Rab27a/b–Slp2-a complex clusters these vesicles at the AMIS. Slp4-a, with Rab3b/Rab8a/Rab27a/b and the t-SNARE syntaxin-3 (Stx3), promotes vesicle fusion at the AMIS. Exocytosis also functions in initial recruitment of the Par3–aPKC complex to the AMIS: Rab8 stimulates Cdc42 loading onto exocytic carriers, possibly through the Cdc42 GEF Tuba. The Cdc42–Par6–aPKC–Par3 complex thus forms at the AMIS, in a step that may co-scaffold the exocyst complex at the AMIS. (c) The AMIS gives rise to the pre-apical patch, where the cell’s apical and basolateral surfaces are distinct, but the apical surfaces are still closely apposed between the two (or more) neighbouring cells. (d) Opening of the lumen results from ion and water transport, and further polarization of the membranes, junctional complexes, cytoskeleton and organelles gives rise to the mature cyst. Figure is modified with permission from ref. .
References
-
- Gassama-Diagne A, et al. Phosphatidylinositol-3,4,5-trisphosphate regulates the formation of the basolateral plasma membrane in epithelial cells. Nat Cell Biol. 2006;8:963–970. - PubMed
-
- Jacob R, Naim HY. Apical membrane proteins are transported in distinct vesicular carriers. Curr Biol. 2001;11:1444–1450. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
