

Conserved regulatory architecture underlies parallel genetic changes and convergent phenotypic evolution
- PMID: 23197832
- PMCID: PMC3529038
- DOI: 10.1073/pnas.1207715109
Conserved regulatory architecture underlies parallel genetic changes and convergent phenotypic evolution
Abstract
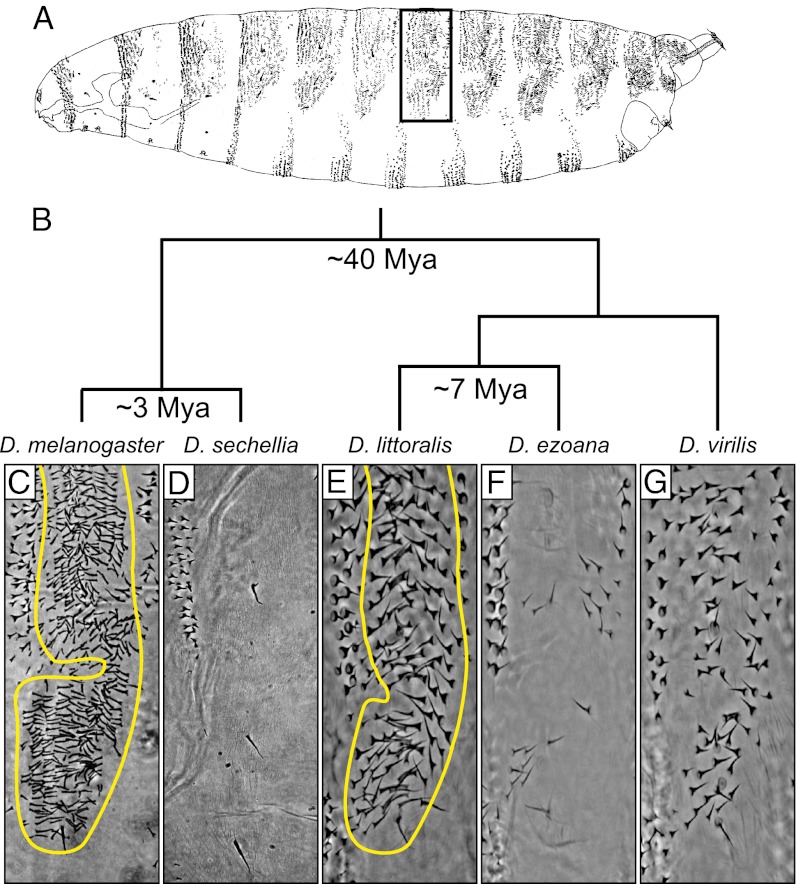
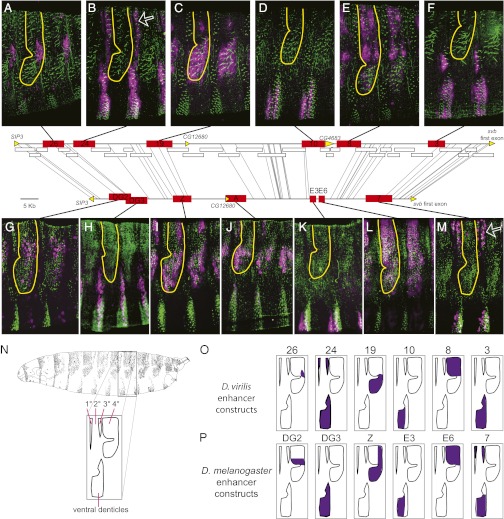
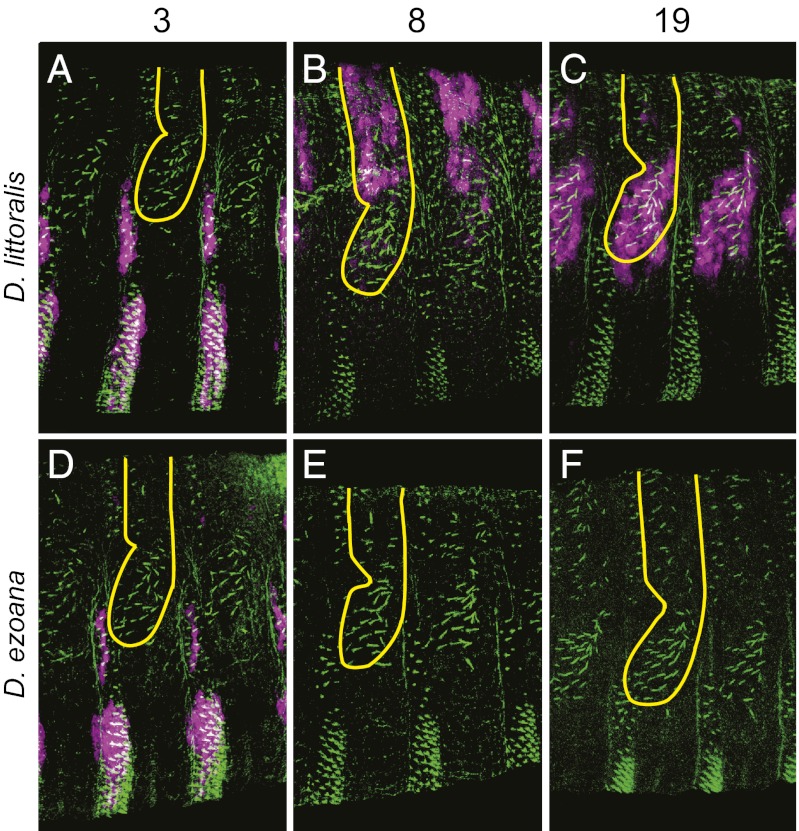
Similar morphological, physiological, and behavioral features have evolved independently in different species, a pattern known as convergence. It is known that morphological convergence can occur through changes in orthologous genes. In some cases of convergence, cis-regulatory changes generate parallel modifications in the expression patterns of orthologous genes. Our understanding of how changes in cis-regulatory regions contribute to convergence is hampered, usually, by a limited understanding of the global cis-regulatory structure of the evolving genes. Here we examine the genetic causes of a case of precise phenotypic convergence between Drosophila sechellia and Drosophila ezoana, species that diverged ~40 Mya. Previous studies revealed that changes in multiple transcriptional enhancers of shavenbaby (svb, a transcript of the ovo locus) caused phenotypic evolution in the D. sechellia lineage. It has also been shown that the convergent phenotype of D. ezoana was likely caused by cis-regulatory evolution of svb. Here we show that the large-scale cis-regulatory architecture of svb is conserved between these Drosophila species. Furthermore, we show that the D. ezoana orthologs of the evolved D. sechellia enhancers have also evolved expression patterns that correlate precisely with the changes in the phenotype. Our results suggest that phenotypic convergence resulted from multiple noncoding changes that occurred in parallel in the D. sechellia and D. ezoana lineages.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Streisfeld MA, Rausher MD. Genetic changes contributing to the parallel evolution of red floral pigmentation among Ipomoea species. New Phytol. 2009;183(3):751–763. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
