

A neuropeptide circuit that coordinates sperm transfer and copulation duration in Drosophila
- PMID: 23197833
- PMCID: PMC3528491
- DOI: 10.1073/pnas.1218246109
A neuropeptide circuit that coordinates sperm transfer and copulation duration in Drosophila
Abstract
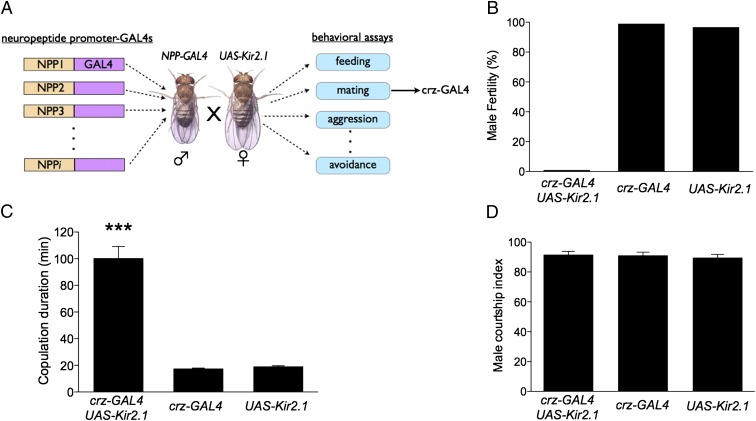
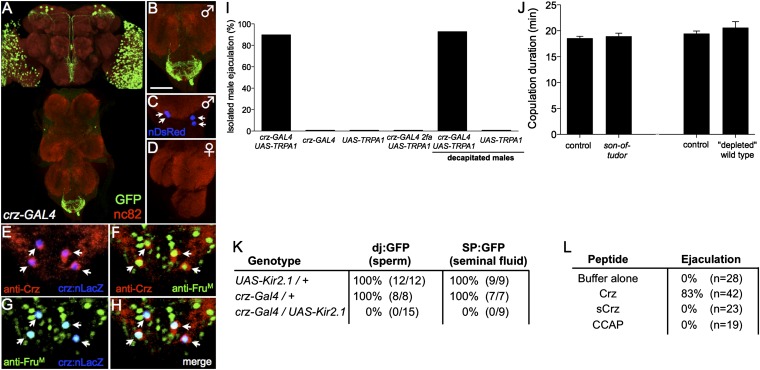
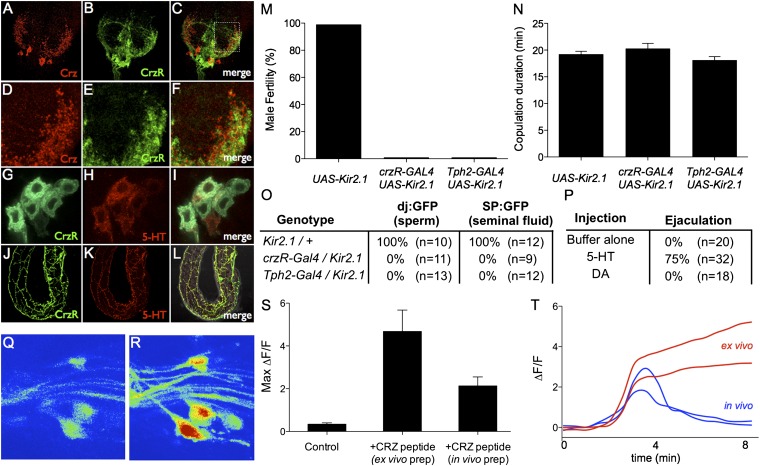
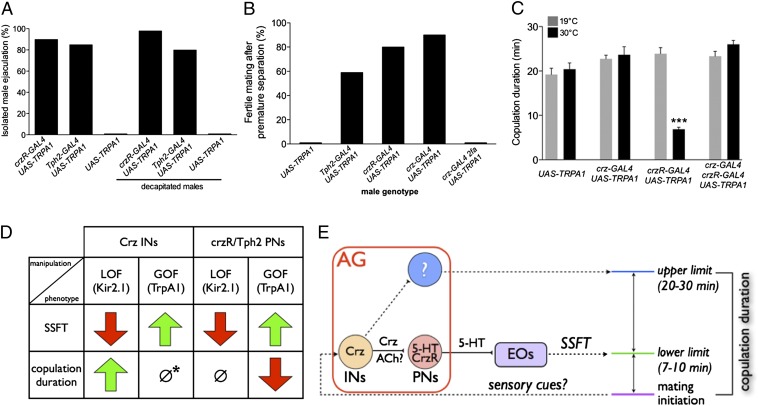
Innate behaviors are often executed in concert with accompanying physiological programs. How this coordination is achieved is poorly understood. Mating behavior and the transfer of sperm and seminal fluid (SSFT) provide a model for understanding how concerted behavioral and physiological programs are coordinated. Here we identify a male-specific neural pathway that coordinates the timing of SSFT with the duration of copulation behavior in Drosophila. Silencing four abdominal ganglion (AG) interneurons (INs) that contain the neuropeptide corazonin (Crz) both blocked SSFT and substantially lengthened copulation duration. Activating these Crz INs caused rapid ejaculation in isolated males, a phenotype mimicked by injection of Crz peptide. Crz promotes SSFT by activating serotonergic (5-HT) projection neurons (PNs) that innervate the accessory glands. Activation of these PNs in copulo caused premature SSFT and also shortened copulation duration. However, mating terminated normally when these PNs were silenced, indicating that SSFT is not required for appropriate copulation duration. Thus, the lengthened copulation duration phenotype caused by silencing Crz INs is independent of the block to SSFT. We conclude that four Crz INs independently control SSFT and copulation duration, thereby coupling the timing of these two processes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wolfner MF. Tokens of love: functions and regulation of Drosophila male accessory gland products. Insect Biochem Mol Biol. 1997;27(3):179–192. - PubMed
-
- Bretman A, Lawniczak MK, Boone J, Chapman T. A mating plug protein reduces early female remating in Drosophila melanogaster. J Insect Physiol. 2010;56(1):107–113. - PubMed
-
- Gillott C. Male accessory gland secretions: Modulators of female reproductive physiology and behavior. Annu Rev Entomol. 2003;48:163–184. - PubMed
-
- Lung O, Wolfner MF. Identification and characterization of the major Drosophila melanogaster mating plug protein. Insect Biochem Mol Biol. 2001;31(6-7):543–551. - PubMed
-
- Gilchrist AS, Partridge L. Why it is difficult to model sperm displacement in Drosophila melanogaster: The relation between sperm transfer and copulation duration. Evolution. 2000;54(2):534–542. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
