

Bradykinetic alcohol dehydrogenases make yeast fitter for growth in the presence of allyl alcohol
- PMID: 23200945
- PMCID: PMC3596495
- DOI: 10.1016/j.cbi.2012.11.010
Bradykinetic alcohol dehydrogenases make yeast fitter for growth in the presence of allyl alcohol
Abstract
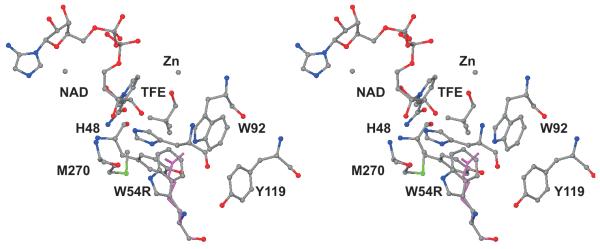
Previous studies showed that fitter yeast (Saccharomyces cerevisiae) that can grow by fermenting glucose in the presence of allyl alcohol, which is oxidized by alcohol dehydrogenase I (ADH1) to toxic acrolein, had mutations in the ADH1 gene that led to decreased ADH activity. These yeast may grow more slowly due to slower reduction of acetaldehyde and a higher NADH/NAD(+) ratio, which should decrease the oxidation of allyl alcohol. We determined steady-state kinetic constants for three yeast ADHs with new site-directed substitutions and examined the correlation between catalytic efficiency and growth on selective media of yeast expressing six different ADHs. The H15R substitution (a test for electrostatic effects) is on the surface of ADH and has small effects on the kinetics. The H44R substitution (affecting interactions with the coenzyme pyrophosphate) was previously shown to decrease affinity for coenzymes 2-4-fold and turnover numbers (V/Et) by 4-6-fold. The W82R substitution is distant from the active site, but decreases turnover numbers by 5-6-fold, perhaps by effects on protein dynamics. The E67Q substitution near the catalytic zinc was shown previously to increase the Michaelis constant for acetaldehyde and to decrease turnover for ethanol oxidation. The W54R substitution, in the substrate binding site, increases kinetic constants (Ks, by >10-fold) while decreasing turnover numbers by 2-7-fold. Growth of yeast expressing the different ADHs on YPD plates (yeast extract, peptone and dextrose) plus antimycin to require fermentation, was positively correlated with the log of catalytic efficiency for the sequential bi reaction (V1/KiaKb=KeqV2/KpKiq, varying over 4 orders of magnitude, adjusted for different levels of ADH expression) in the order: WT≈H15R>H44R>W82R>E67Q>W54R. Growth on YPD plus 10mM allyl alcohol was inversely correlated with catalytic efficiency. The fitter yeast are "bradytrophs" (slow growing) because the ADHs have decreased catalytic efficiency.
Copyright © 2012 Elsevier Ireland Ltd. All rights reserved.
Figures




Similar articles
-
The Thr45Gly substitution in yeast alcohol dehydrogenase substantially decreases catalysis, alters pH dependencies, and disrupts the proton relay system.Chem Biol Interact. 2021 Nov 1;349:109650. doi: 10.1016/j.cbi.2021.109650. Epub 2021 Sep 13. Chem Biol Interact. 2021. PMID: 34529977 Free PMC article.
-
Substitution of cysteine-153 ligated to the catalytic zinc in yeast alcohol dehydrogenase with aspartic acid and analysis of mechanisms of related medium chain dehydrogenases.Chem Biol Interact. 2019 Apr 1;302:172-182. doi: 10.1016/j.cbi.2019.01.040. Epub 2019 Feb 2. Chem Biol Interact. 2019. PMID: 30721696
-
Substitution of both histidines in the active site of yeast alcohol dehydrogenase 1 exposes underlying pH dependencies.Chem Biol Interact. 2024 May 1;394:110992. doi: 10.1016/j.cbi.2024.110992. Epub 2024 Apr 4. Chem Biol Interact. 2024. PMID: 38579923 Free PMC article.
-
The alcohol dehydrogenases of Saccharomyces cerevisiae: a comprehensive review.FEMS Yeast Res. 2008 Nov;8(7):967-78. doi: 10.1111/j.1567-1364.2008.00387.x. Epub 2008 May 7. FEMS Yeast Res. 2008. PMID: 18479436 Review.
-
Natural alcohol exposure: is ethanol the main substrate for alcohol dehydrogenases in animals?Chem Biol Interact. 2011 May 30;191(1-3):14-25. doi: 10.1016/j.cbi.2011.02.008. Epub 2011 Feb 15. Chem Biol Interact. 2011. PMID: 21329681 Review.
Cited by
-
Unbalance between Pyridine Nucleotide Cofactors in The SOD1 Deficient Yeast Saccharomyces cerevisiae Causes Hypersensitivity to Alcohols and Aldehydes.Int J Mol Sci. 2022 Dec 30;24(1):659. doi: 10.3390/ijms24010659. Int J Mol Sci. 2022. PMID: 36614102 Free PMC article.
-
Yeast alcohol dehydrogenase structure and catalysis.Biochemistry. 2014 Sep 16;53(36):5791-803. doi: 10.1021/bi5006442. Epub 2014 Sep 3. Biochemistry. 2014. PMID: 25157460 Free PMC article.
-
Increased alcohol dehydrogenase 1 activity promotes longevity.Curr Biol. 2023 Mar 27;33(6):1036-1046.e6. doi: 10.1016/j.cub.2023.01.059. Epub 2023 Feb 17. Curr Biol. 2023. PMID: 36805847 Free PMC article.
-
Macromolecular crowding effects on the kinetics of opposing reactions catalyzed by alcohol dehydrogenase.Biochem Biophys Rep. 2021 Feb 20;26:100956. doi: 10.1016/j.bbrep.2021.100956. eCollection 2021 Jul. Biochem Biophys Rep. 2021. PMID: 33665382 Free PMC article.
-
The Alcohol Dehydrogenase Gene Family in Melon (Cucumis melo L.): Bioinformatic Analysis and Expression Patterns.Front Plant Sci. 2016 May 18;7:670. doi: 10.3389/fpls.2016.00670. eCollection 2016. Front Plant Sci. 2016. PMID: 27242871 Free PMC article.
References
-
- Wills C, Phelps J. A technique for the isolation of yeast alcohol dehydrogenase mutants with altered substrate specificity. Arch. Biochem. Biophys. 1975;167:627–637. - PubMed
-
- Wills C. Controlling protein evolution. Fed. Proc. 1976;35:2098–2101. - PubMed
-
- Wills C. Production of yeast alcohol dehydrogenase isoenzymes by selection. Nature. 1976;261:26–29. - PubMed
-
- Wills C, Kratofil P, Martin T. Functional mutants of yeast alcohol dehydrogenase. In: Hollaender A, editor. Genetic Engineering of Microorganisms for Chemicals. Plenum; New York: 1981. pp. 305–329.
-
- Wills C, Phelps J. Functional mutants of yeast alcohol dehydrogenase affecting kinetics, cellular redox balance, and electrophoretic mobility. Biochem. Genet. 1978;16:415–432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
