

Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions
- PMID: 23201972
- PMCID: PMC4136922
- DOI: 10.1038/nn.3275
Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions
Abstract
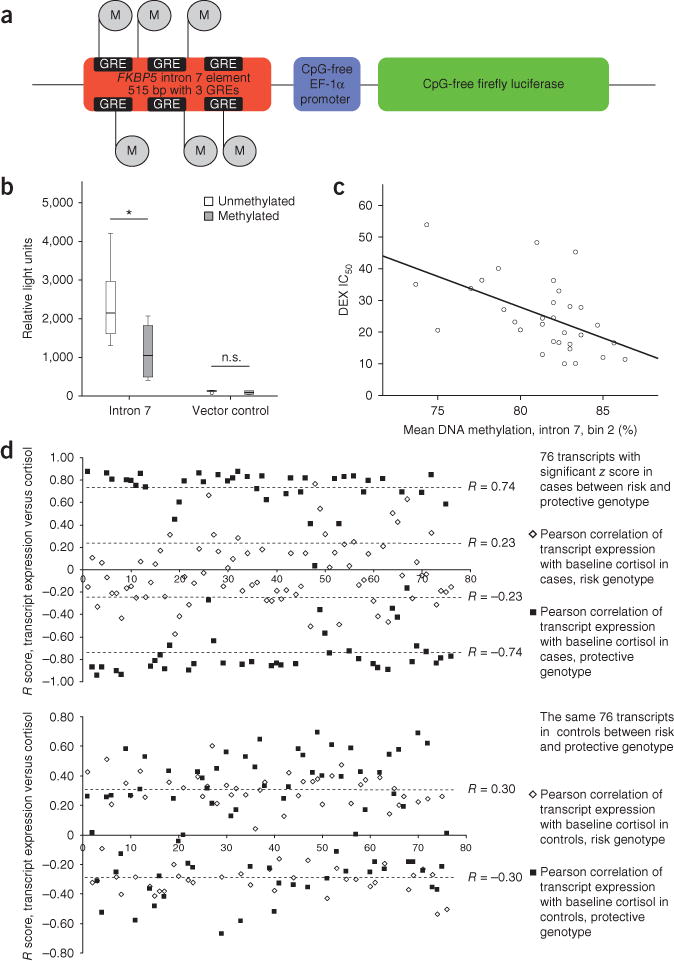
Although the fact that genetic predisposition and environmental exposures interact to shape development and function of the human brain and, ultimately, the risk of psychiatric disorders has drawn wide interest, the corresponding molecular mechanisms have not yet been elucidated. We found that a functional polymorphism altering chromatin interaction between the transcription start site and long-range enhancers in the FK506 binding protein 5 (FKBP5) gene, an important regulator of the stress hormone system, increased the risk of developing stress-related psychiatric disorders in adulthood by allele-specific, childhood trauma-dependent DNA demethylation in functional glucocorticoid response elements of FKBP5. This demethylation was linked to increased stress-dependent gene transcription followed by a long-term dysregulation of the stress hormone system and a global effect on the function of immune cells and brain areas associated with stress regulation. This identification of molecular mechanisms of genotype-directed long-term environmental reactivity will be useful for designing more effective treatment strategies for stress-related disorders.
Conflict of interest statement
Figures






Comment in
-
Neurogenetics: Trauma and stress, from child to adult.Nat Rev Genet. 2013 Feb;14(2):77. doi: 10.1038/nrg3406. Epub 2012 Dec 19. Nat Rev Genet. 2013. PMID: 23249871 No abstract available.
-
Neurogenetics: Trauma and stress, from child to adult.Nat Rev Neurosci. 2013 Feb;14(2):77. doi: 10.1038/nrn3425. Epub 2012 Dec 19. Nat Rev Neurosci. 2013. PMID: 23249990 No abstract available.
-
How do environments talk to genes?Nat Neurosci. 2013 Jan;16(1):2-4. doi: 10.1038/nn.3286. Nat Neurosci. 2013. PMID: 23257925 No abstract available.
References
-
- Caspi A, Moffitt TE. Gene-environment interactions in psychiatry: joining forces with neuroscience. Nat Rev Neurosci. 2006;7:583–590. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
