

The SWI2/SNF2 chromatin remodeling ATPase BRAHMA represses abscisic acid responses in the absence of the stress stimulus in Arabidopsis
- PMID: 23209114
- PMCID: PMC3556964
- DOI: 10.1105/tpc.112.105114
The SWI2/SNF2 chromatin remodeling ATPase BRAHMA represses abscisic acid responses in the absence of the stress stimulus in Arabidopsis
Abstract
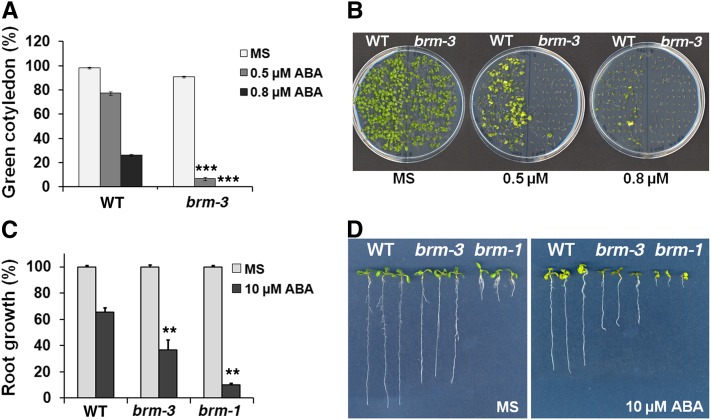
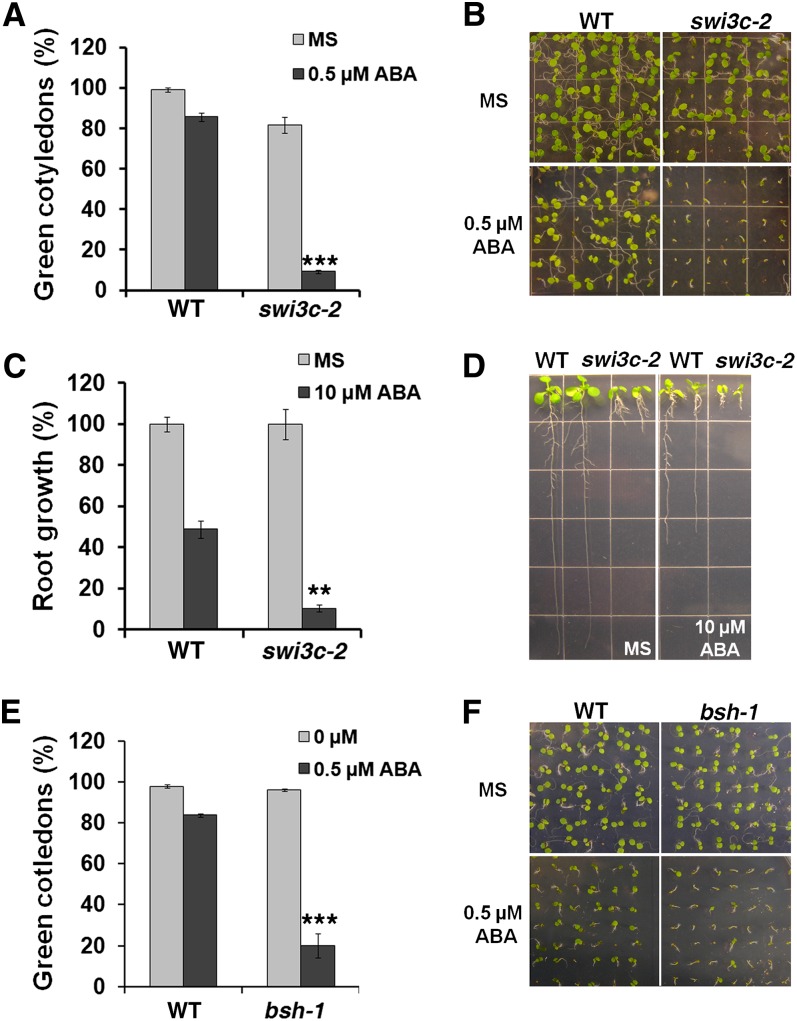
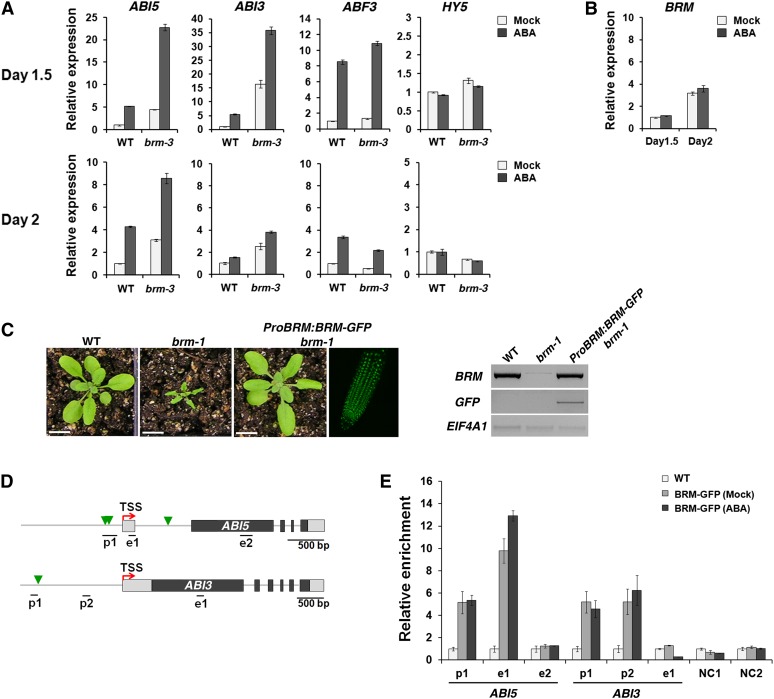
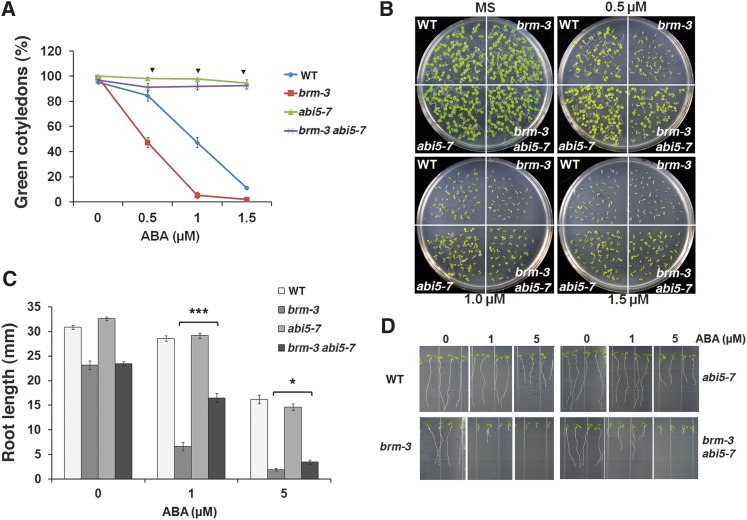
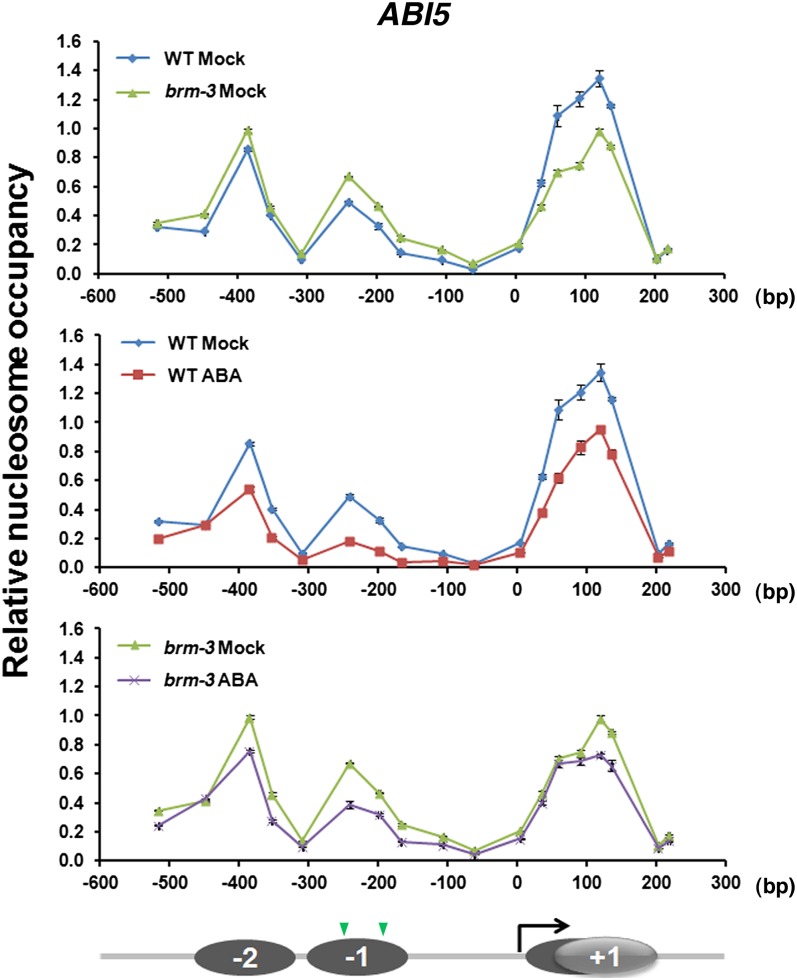
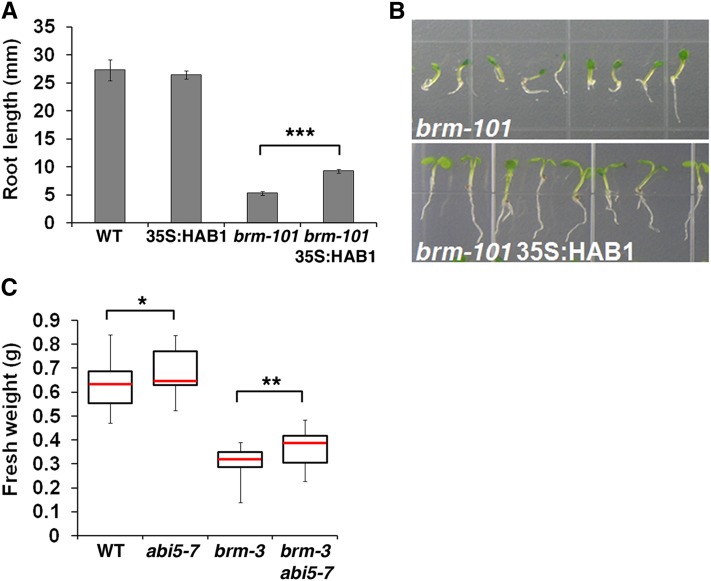
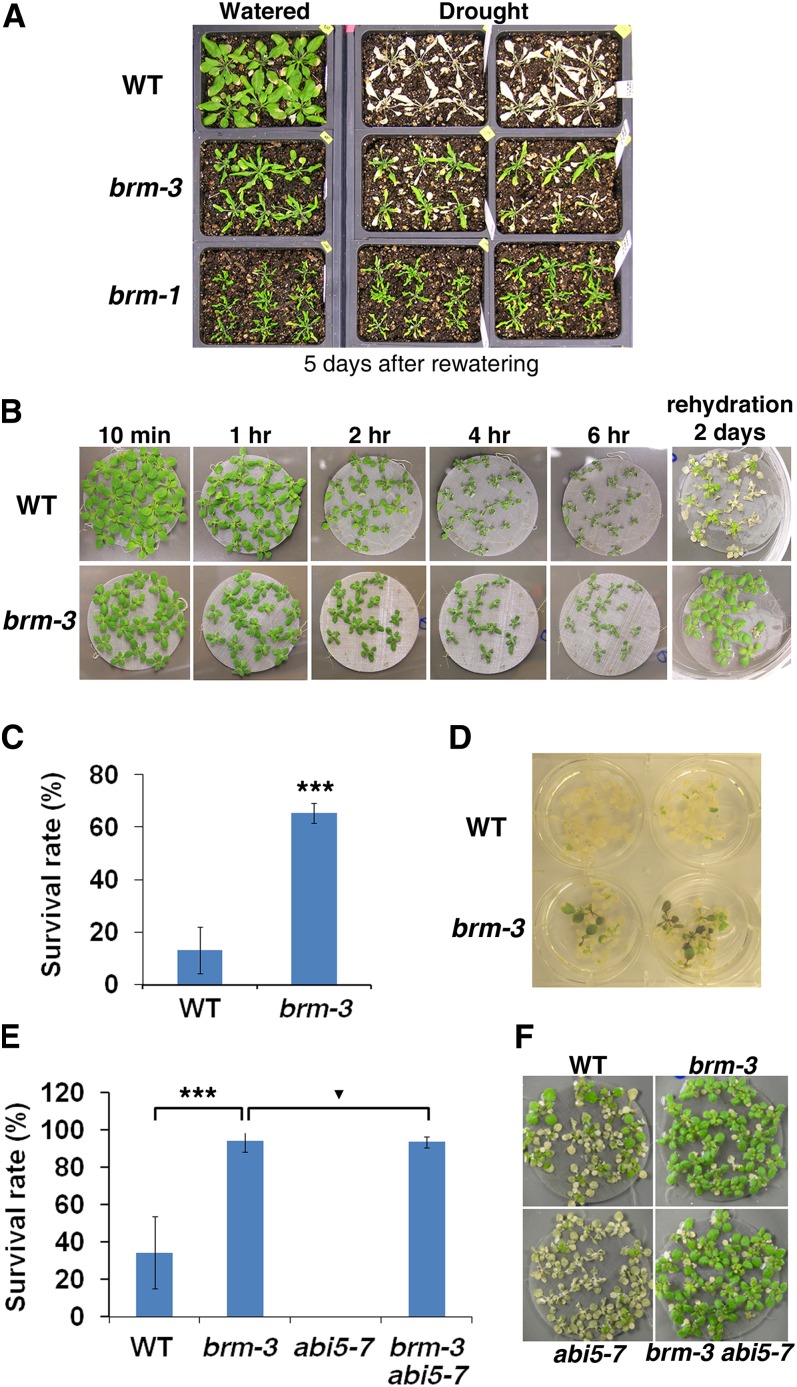
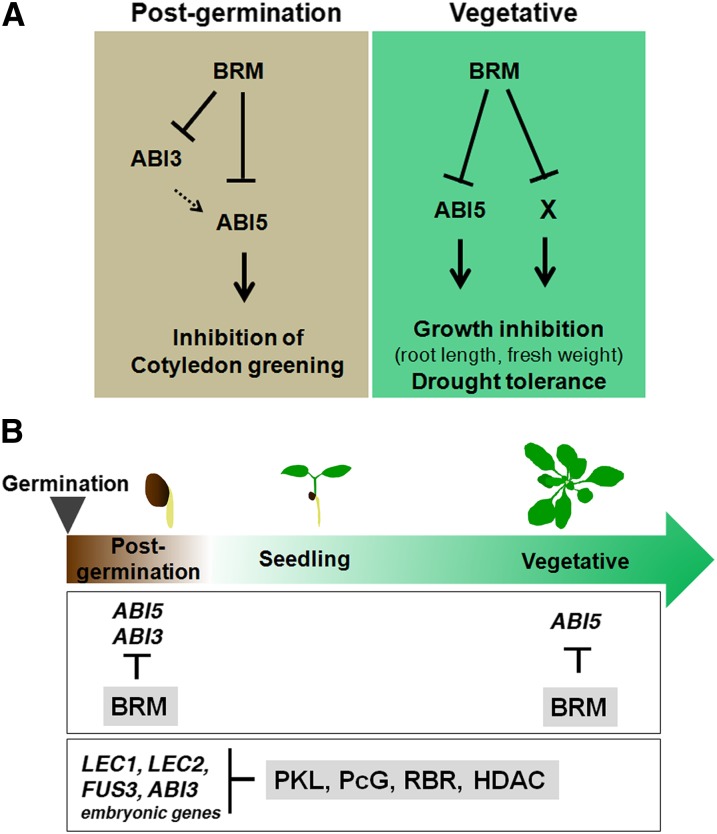
The survival of plants as sessile organisms depends on their ability to cope with environmental challenges. Of key importance in this regard is the phytohormone abscisic acid (ABA). ABA not only promotes seed dormancy but also triggers growth arrest in postgermination embryos that encounter water stress. This is accompanied by increased desiccation tolerance. Postgermination ABA responses in Arabidopsis thaliana are mediated in large part by the ABA-induced basic domain/leucine zipper transcription factor ABA INSENSITIVE5 (ABI5). Here, we show that loss of function of the SWI2/SNF2 chromatin remodeling ATPase BRAHMA (BRM) causes ABA hypersensitivity during postgermination growth arrest. ABI5 expression was derepressed in brm mutants in the absence of exogenous ABA and accumulated to high levels upon ABA sensing. This effect was likely direct; chromatin immunoprecipitation revealed BRM binding to the ABI5 locus. Moreover, loss of BRM activity led to destabilization of a nucleosome likely to repress ABI5 transcription. Finally, the abi5 null mutant was epistatic to BRM in postgermination growth arrest. In addition, vegetative growth defects typical of brm mutants in the absence of ABA treatment could be partially overcome by reduction of ABA responses, and brm mutants displayed increased drought tolerance. We propose a role for BRM in the balance between growth or stress responses.
Figures
References
-
- Alon U. (2007). Network motifs: Theory and experimental approaches. Nat. Rev. Genet. 8: 450–461 - PubMed
-
- Archacki R., Sarnowski T.J., Halibart-Puzio J., Brzeska K., Buszewicz D., Prymakowska-Bosak M., Koncz C., Jerzmanowski A. (2009). Genetic analysis of functional redundancy of BRM ATPase and ATSWI3C subunits of Arabidopsis SWI/SNF chromatin remodelling complexes. Planta 229: 1281–1292 - PubMed
-
- Battisti D.S., Naylor R.L. (2009). Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323: 240–244 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
