

Epigenetics and bacterial infections
- PMID: 23209181
- PMCID: PMC3543073
- DOI: 10.1101/cshperspect.a010272
Epigenetics and bacterial infections
Abstract
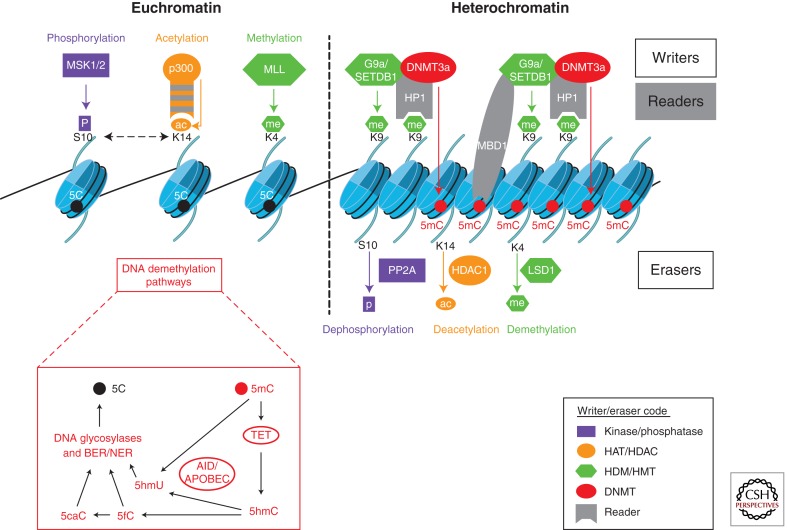
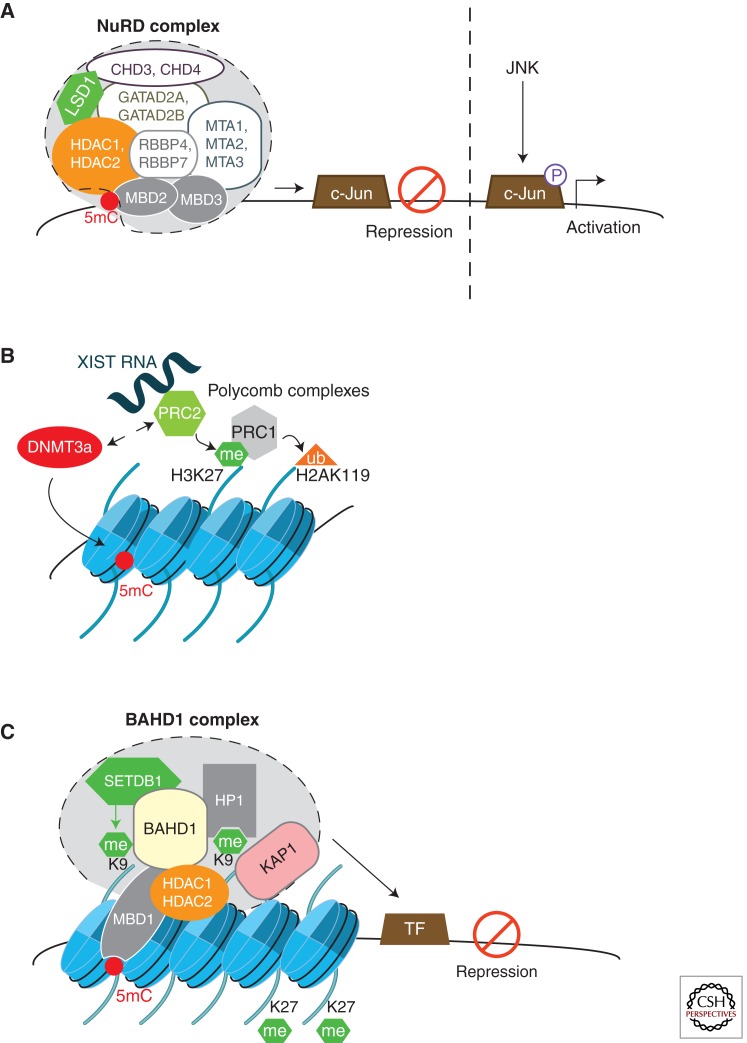
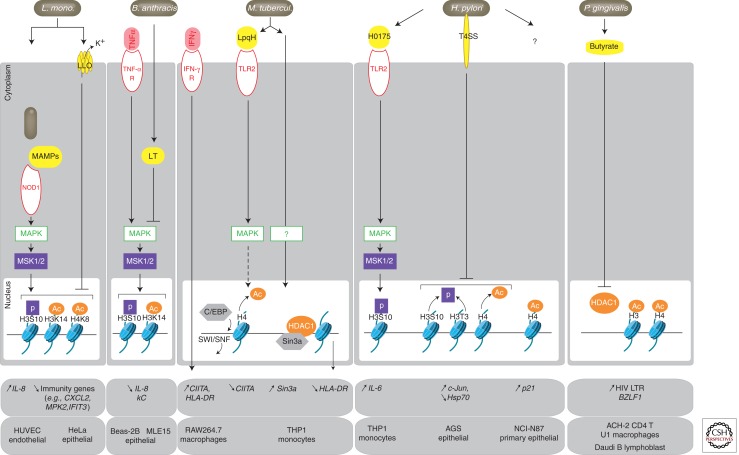
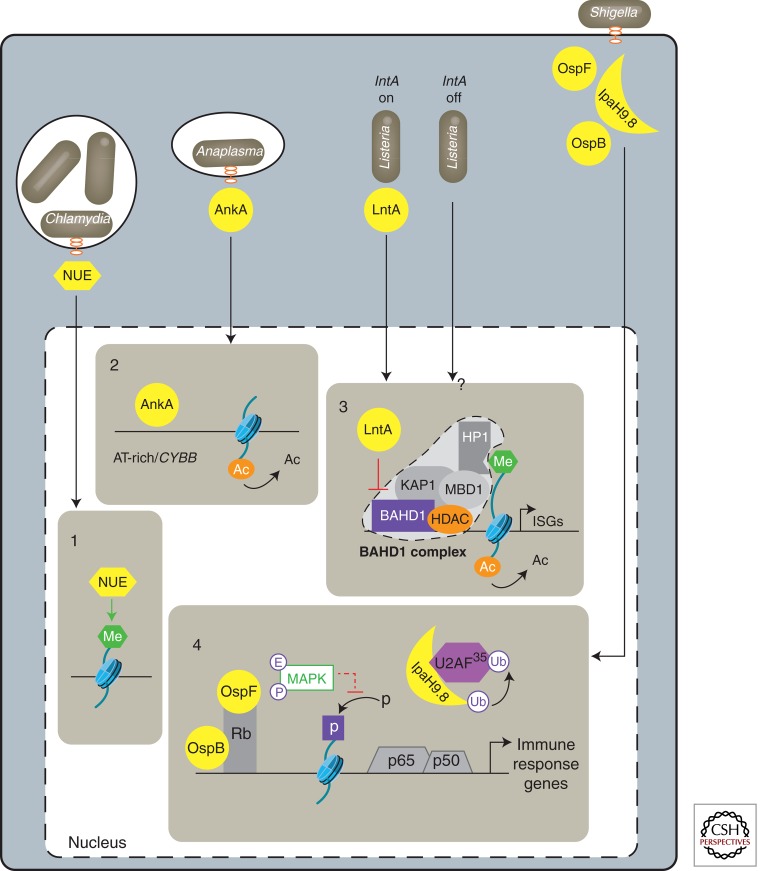
Epigenetic mechanisms regulate expression of the genome to generate various cell types during development or orchestrate cellular responses to external stimuli. Recent studies highlight that bacteria can affect the chromatin structure and transcriptional program of host cells by influencing diverse epigenetic factors (i.e., histone modifications, DNA methylation, chromatin-associated complexes, noncoding RNAs, and RNA splicing factors). In this article, we first review the molecular bases of the epigenetic language and then describe the current state of research regarding how bacteria can alter epigenetic marks and machineries. Bacterial-induced epigenetic deregulations may affect host cell function either to promote host defense or to allow pathogen persistence. Thus, pathogenic bacteria can be considered as potential epimutagens able to reshape the epigenome. Their effects might generate specific, long-lasting imprints on host cells, leading to a memory of infection that influences immunity and might be at the origin of unexplained diseases.
Figures
Similar articles
-
Epigenomics in stress tolerance of plants under the climate change.Mol Biol Rep. 2023 Jul;50(7):6201-6216. doi: 10.1007/s11033-023-08539-6. Epub 2023 Jun 9. Mol Biol Rep. 2023. PMID: 37294468 Review.
-
Histone modifications and chromatin remodeling during bacterial infections.Cell Host Microbe. 2008 Aug 14;4(2):100-9. doi: 10.1016/j.chom.2008.07.009. Cell Host Microbe. 2008. PMID: 18692770 Review.
-
The role of epigenetics in tuberculosis infection.Epigenomics. 2016 Apr;8(4):537-49. doi: 10.2217/epi.16.1. Epub 2016 Apr 1. Epigenomics. 2016. PMID: 27035266 Review.
-
Involvement of noncoding RNAs in epigenetic modifications of esophageal cancer.Biomed Pharmacother. 2019 Sep;117:109192. doi: 10.1016/j.biopha.2019.109192. Epub 2019 Jul 11. Biomed Pharmacother. 2019. PMID: 31387188 Review.
-
Epitranscriptomics and epigenetics: two sides of the same coin?Clin Epigenetics. 2024 Sep 3;16(1):121. doi: 10.1186/s13148-024-01729-4. Clin Epigenetics. 2024. PMID: 39252109 Free PMC article. Review.
Cited by
-
Mycobacterium tuberculosis and host interactions in the manifestation of tuberculosis.J Clin Tuberc Other Mycobact Dis. 2024 Jun 14;36:100458. doi: 10.1016/j.jctube.2024.100458. eCollection 2024 Aug. J Clin Tuberc Other Mycobact Dis. 2024. PMID: 38983441 Free PMC article. Review.
-
Mapping epigenetic changes to the host cell genome induced by Burkholderia pseudomallei reveals pathogen-specific and pathogen-generic signatures of infection.Sci Rep. 2016 Aug 3;6:30861. doi: 10.1038/srep30861. Sci Rep. 2016. PMID: 27484700 Free PMC article.
-
Circulating Inflammatory-Associated Proteins in the First Month of Life and Cognitive Impairment at Age 10 Years in Children Born Extremely Preterm.J Pediatr. 2017 Jan;180:116-123.e1. doi: 10.1016/j.jpeds.2016.09.054. Epub 2016 Oct 24. J Pediatr. 2017. PMID: 27788929 Free PMC article.
-
Bovine neonatal microbiome origins: a review of proposed microbial community presence from conception to colostrum.Transl Anim Sci. 2023 May 27;7(1):txad057. doi: 10.1093/tas/txad057. eCollection 2023 Jan. Transl Anim Sci. 2023. PMID: 37334245 Free PMC article.
-
Epigenetic dysfunctional diseases and therapy for infection and inflammation.Biochim Biophys Acta Mol Basis Dis. 2017 Feb;1863(2):518-528. doi: 10.1016/j.bbadis.2016.11.030. Epub 2016 Dec 3. Biochim Biophys Acta Mol Basis Dis. 2017. PMID: 27919711 Free PMC article. Review.
References
-
- Aguilera C, Nakagawa K, Sancho R, Chakraborty A, Hendrich B, Behrens A 2011. c-Jun N-terminal phosphorylation antagonises recruitment of the Mbd3/NuRD repressor complex. Nature 469: 231–235 - PubMed
-
- Allemand E, Batsche E, Muchardt C 2008. Splicing, transcription, and chromatin: A menage a trois. Curr Opin Genet Dev 18: 145–151 - PubMed
-
- Allo M, Buggiano V, Fededa JP, Petrillo E, Schor I, de la Mata M, Agirre E, Plass M, Eyras E, Elela SA, et al. 2009. Control of alternative splicing through siRNA-mediated transcriptional gene silencing. Nat Struct Mol Biol 16: 717–724 - PubMed
-
- Ando T, Yoshida T, Enomoto S, Asada K, Tatematsu M, Ichinose M, Sugiyama T, Ushijima T 2009. DNA methylation of microRNA genes in gastric mucosae of gastric cancer patients: Its possible involvement in the formation of epigenetic field defect. Int J Cancer 124: 2367–2374 - PubMed
-
- Aravind L, Abhiman S, Iyer LM 2011. Natural history of the eukaryotic chromatin protein methylation system. Prog Mol Biol Transl Sci 101: 105–176 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical