

KIF14 negatively regulates Rap1a-Radil signaling during breast cancer progression
- PMID: 23209302
- PMCID: PMC3518219
- DOI: 10.1083/jcb.201206051
KIF14 negatively regulates Rap1a-Radil signaling during breast cancer progression
Abstract
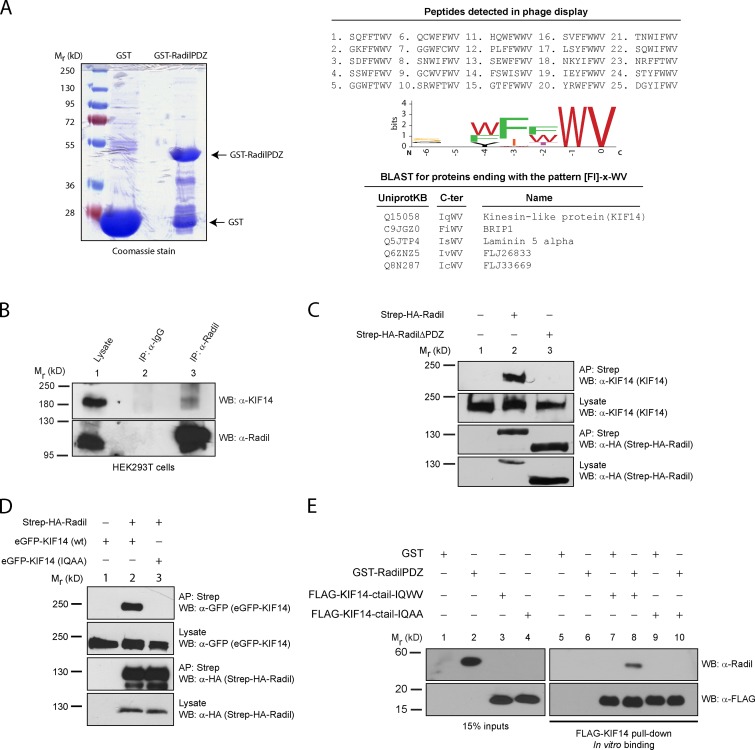
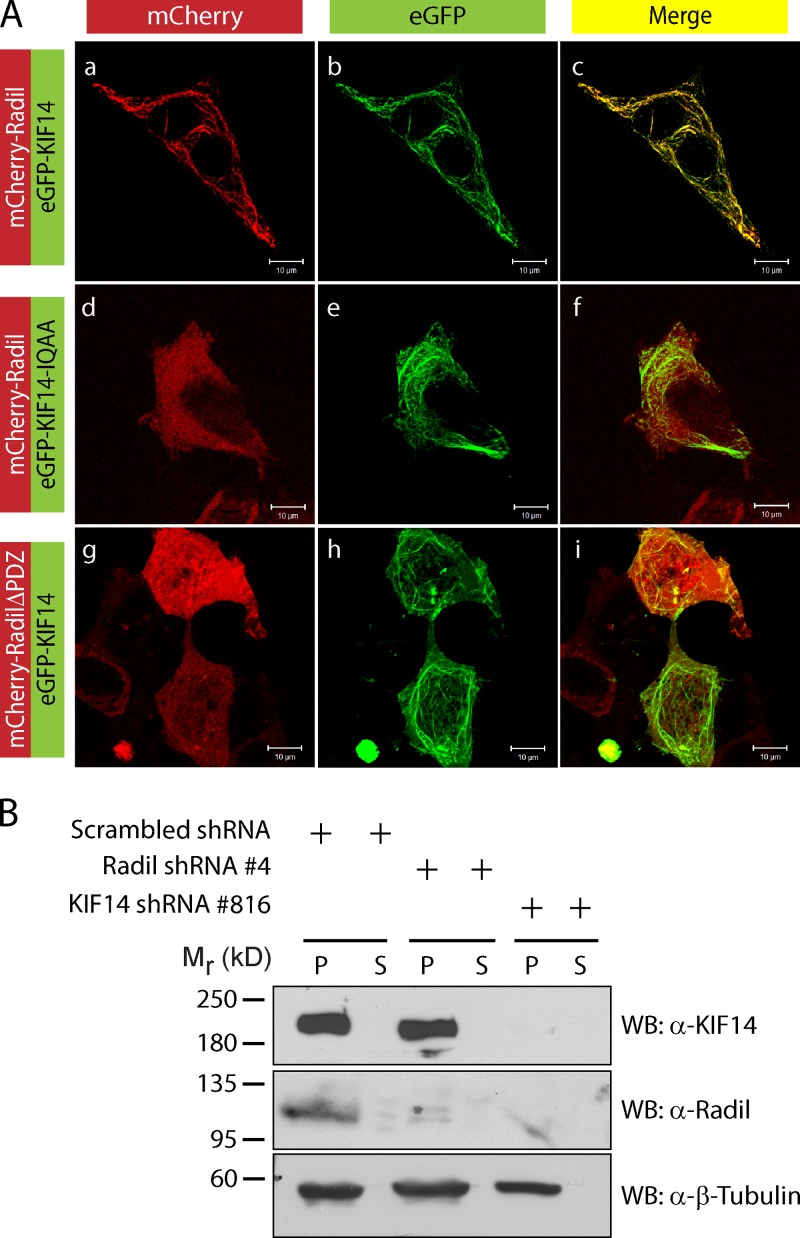
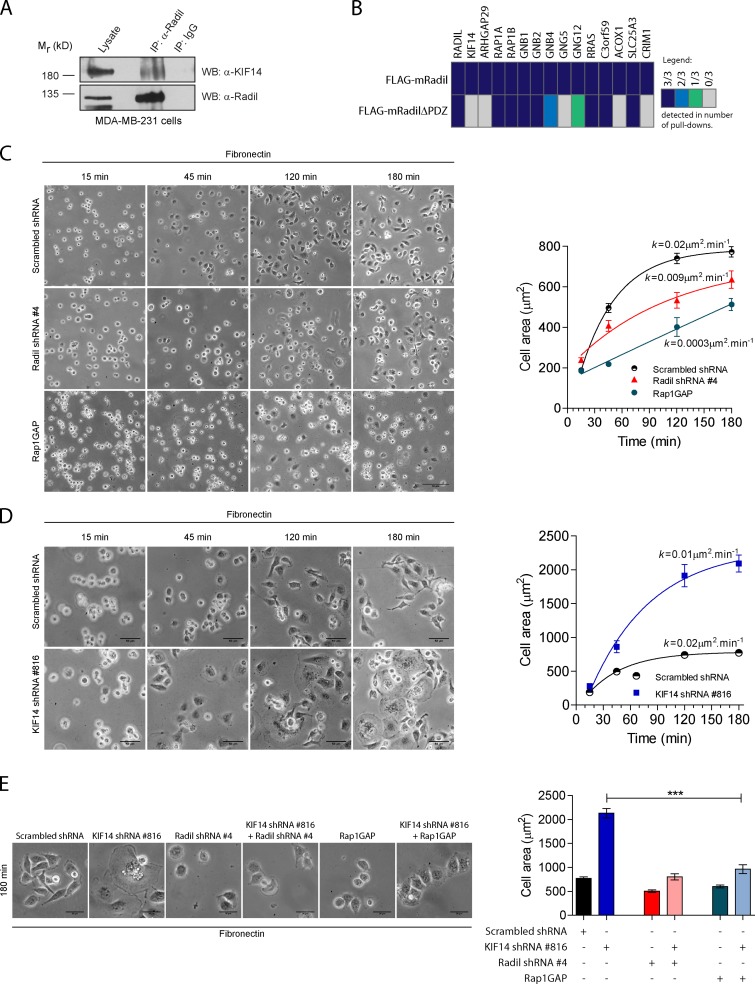
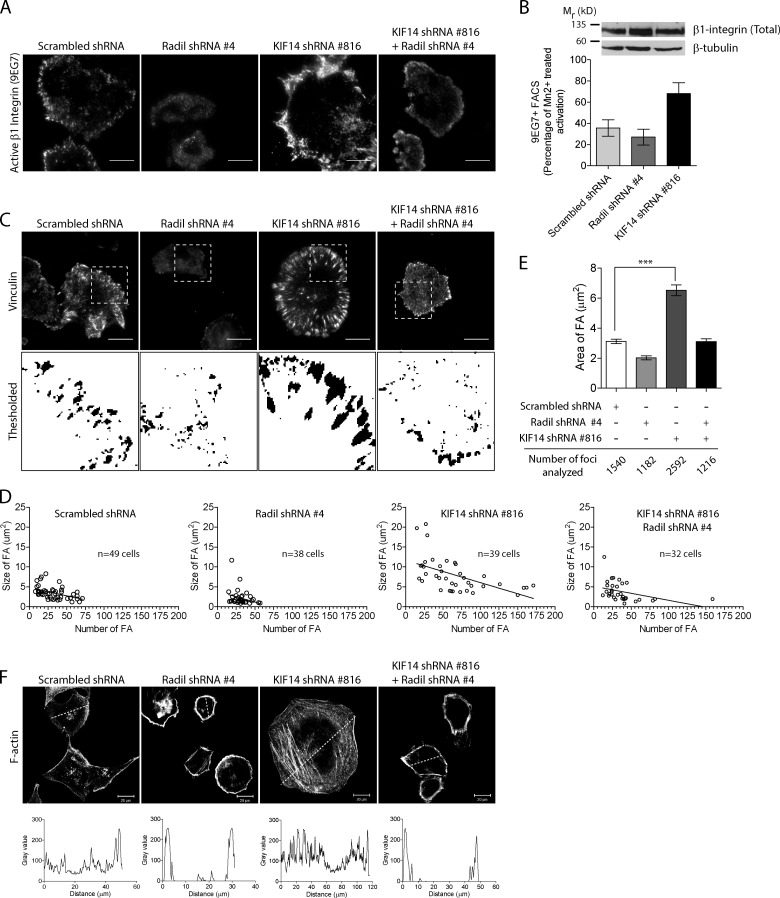
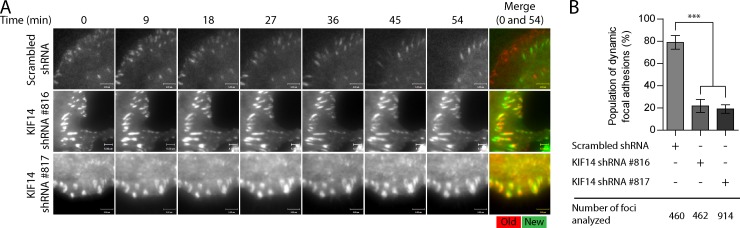
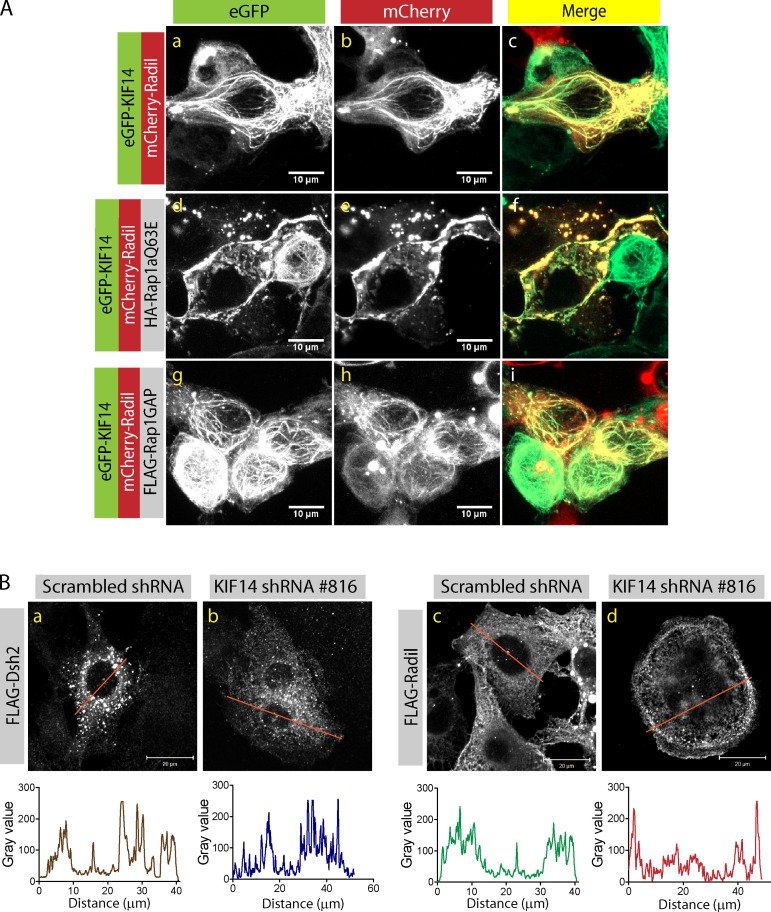
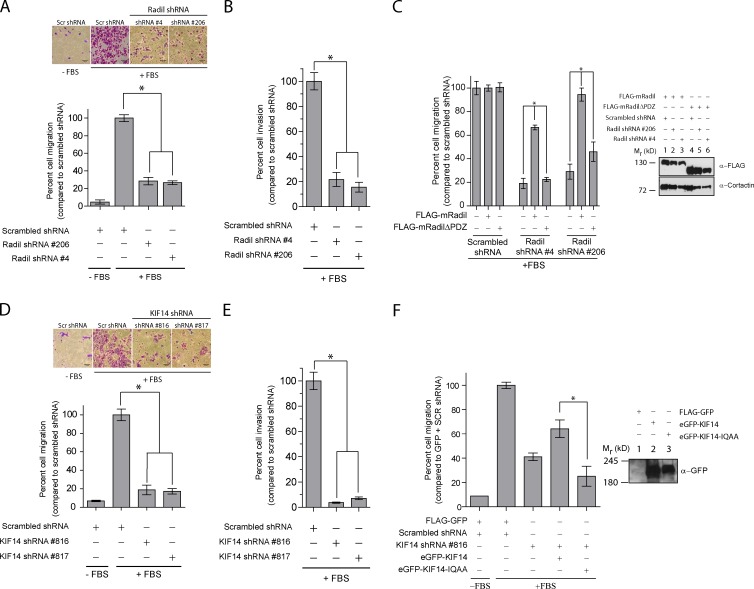
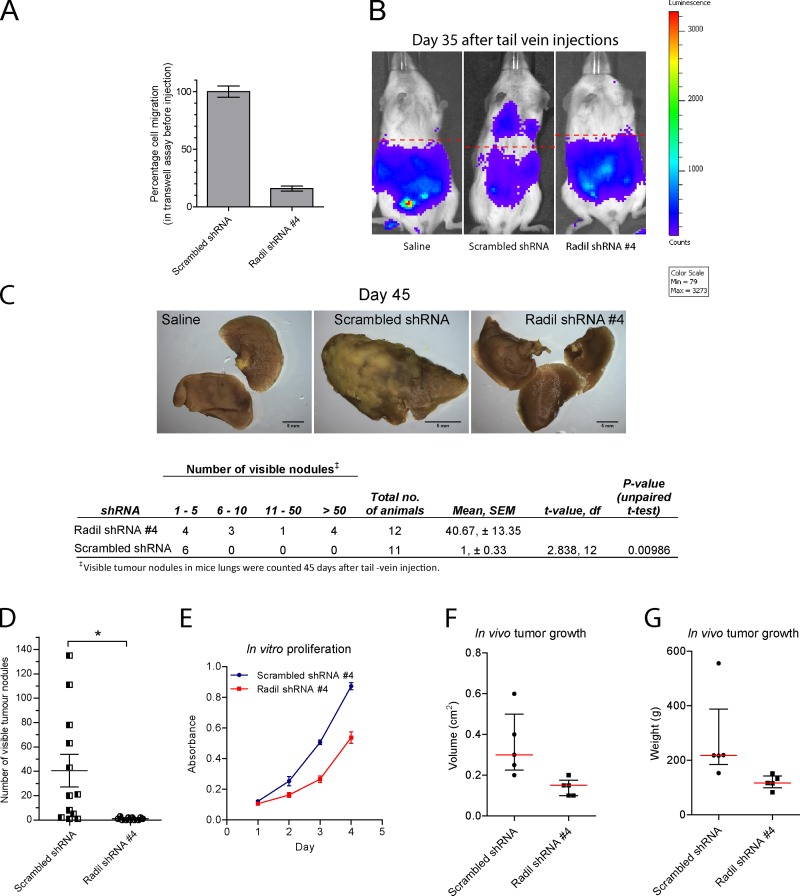
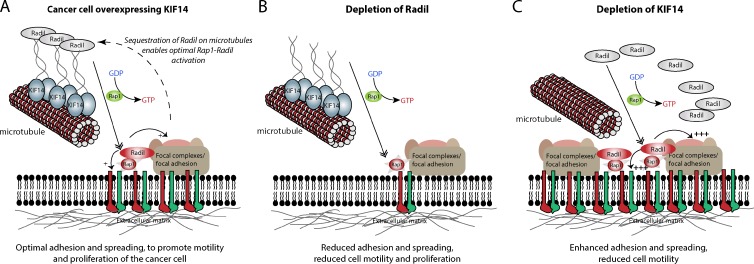
The small GTPase Rap1 regulates inside-out integrin activation and thereby influences cell adhesion, migration, and polarity. Several Rap1 effectors have been described to mediate the cellular effects of Rap1 in a context-dependent manner. Radil is emerging as an important Rap effector implicated in cell spreading and migration, but the molecular mechanisms underlying its functions are unclear. We report here that the kinesin KIF14 associates with the PDZ domain of Radil and negatively regulates Rap1-mediated inside-out integrin activation by tethering Radil on microtubules. The depletion of KIF14 led to increased cell spreading, altered focal adhesion dynamics, and inhibition of cell migration and invasion. We also show that Radil is important for breast cancer cell proliferation and for metastasis in mice. Our findings provide evidence that the concurrent up-regulation of Rap1 activity and increased KIF14 levels in several cancers is needed to reach optimal levels of Rap1-Radil signaling, integrin activation, and cell-matrix adhesiveness required for tumor progression.
Figures
References
-
- Alley M.C., Scudiero D.A., Monks A., Hursey M.L., Czerwinski M.J., Fine D.L., Abbott B.J., Mayo J.G., Shoemaker R.H., Boyd M.R. 1988. Feasibility of drug screening with panels of human tumor cell lines using a microculture tetrazolium assay. Cancer Res. 48:589–601 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
