

Bub1 kinase activity drives error correction and mitotic checkpoint control but not tumor suppression
- PMID: 23209306
- PMCID: PMC3518220
- DOI: 10.1083/jcb.201205115
Bub1 kinase activity drives error correction and mitotic checkpoint control but not tumor suppression
Abstract
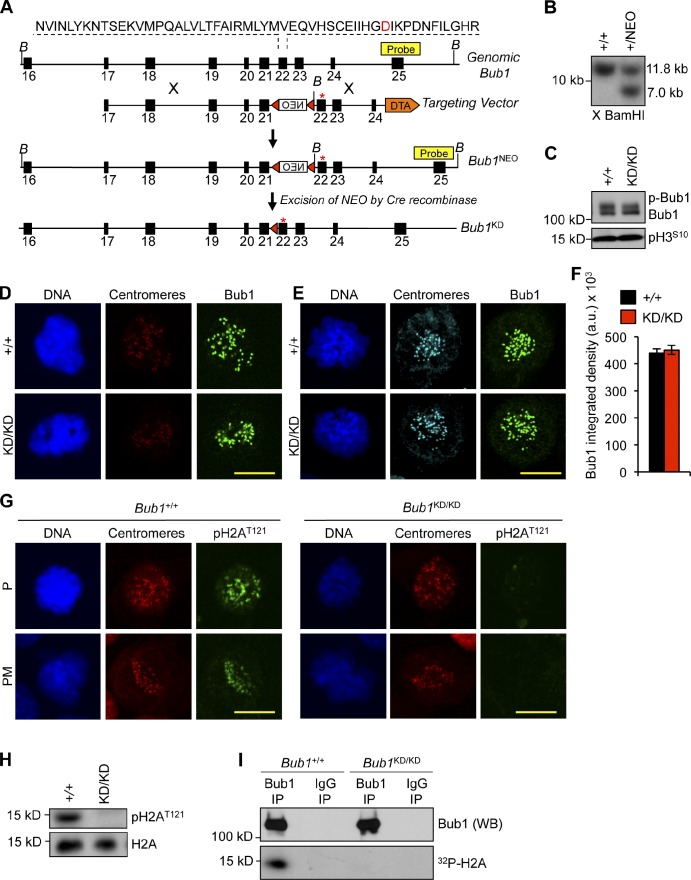
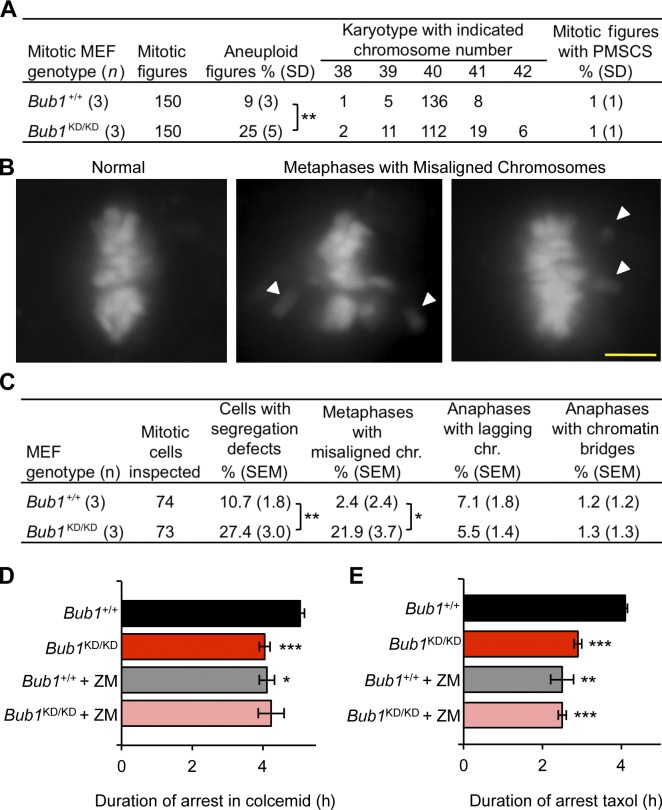
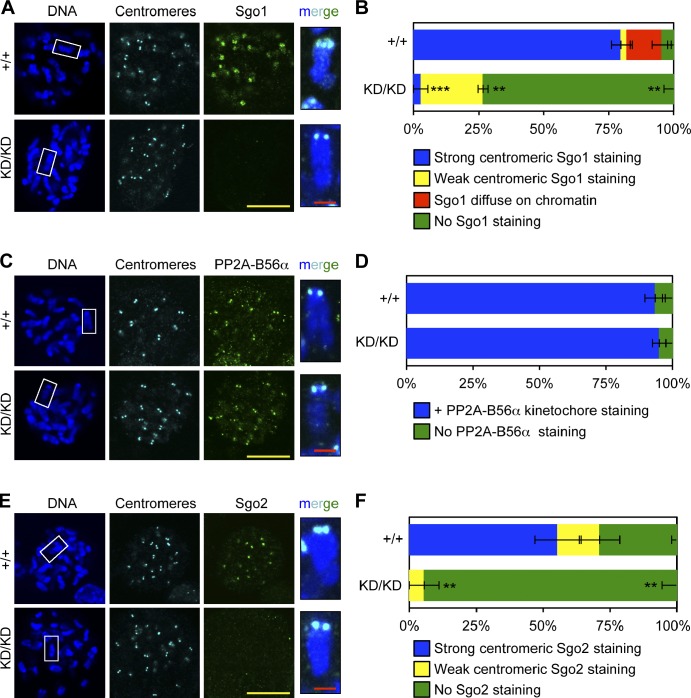
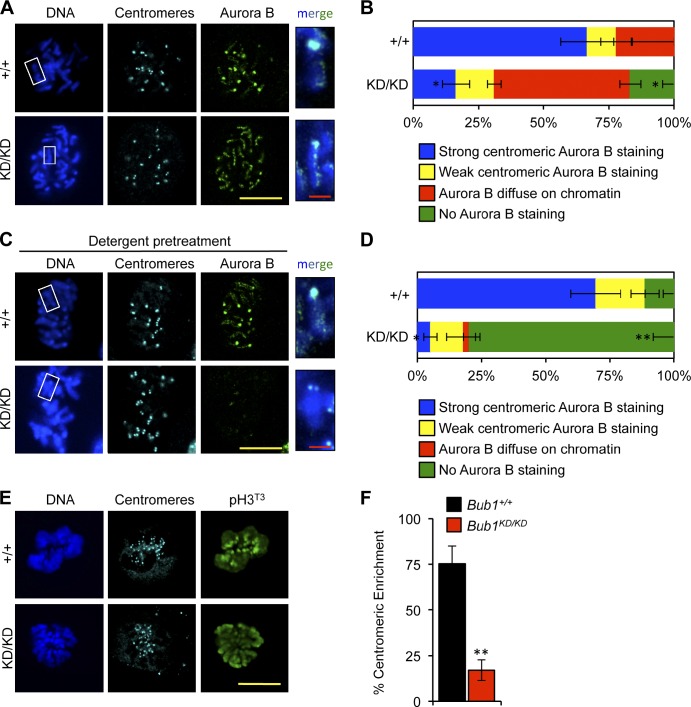
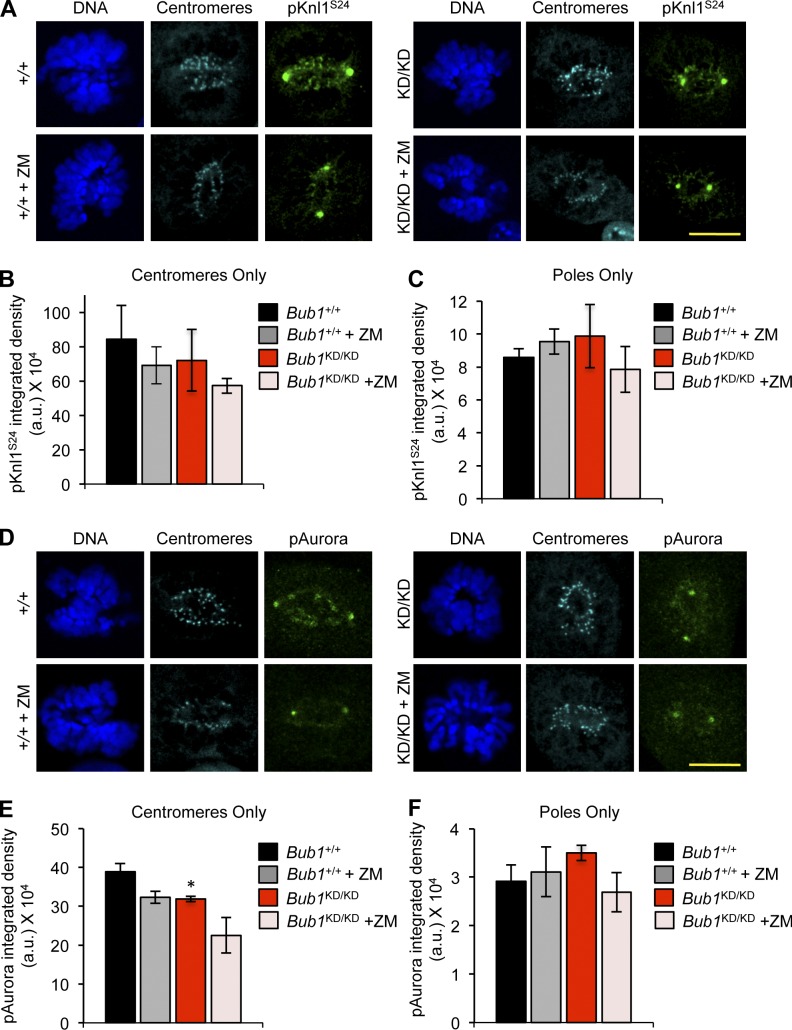
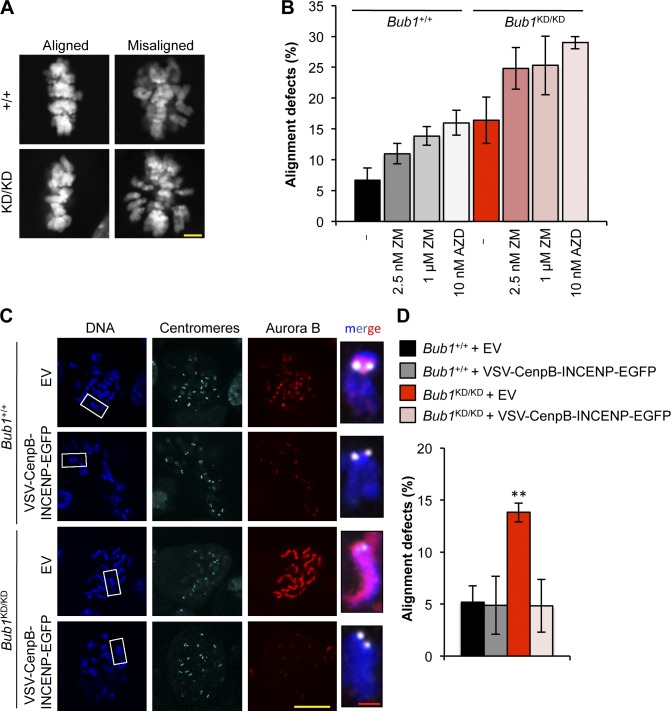
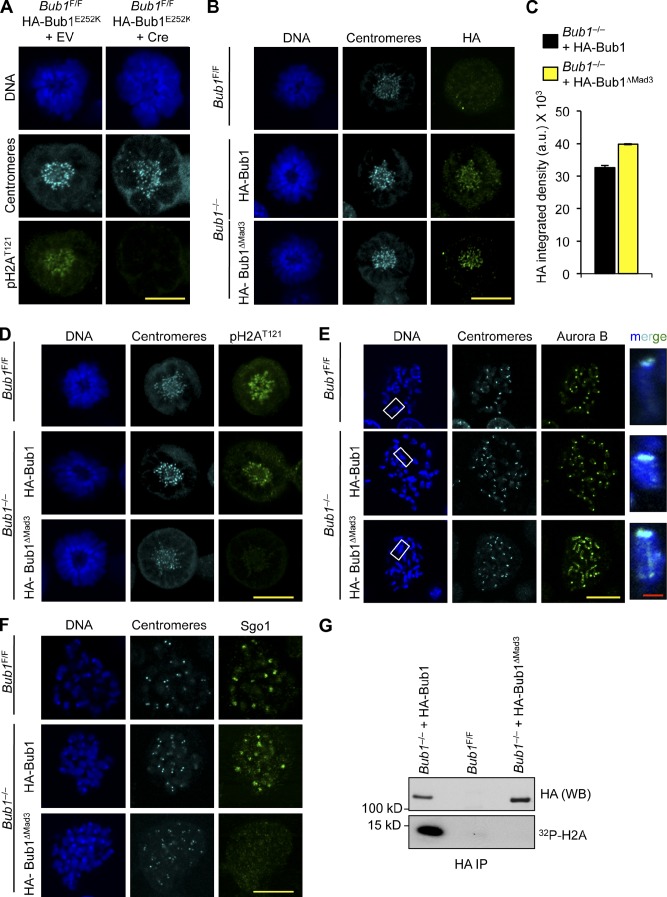
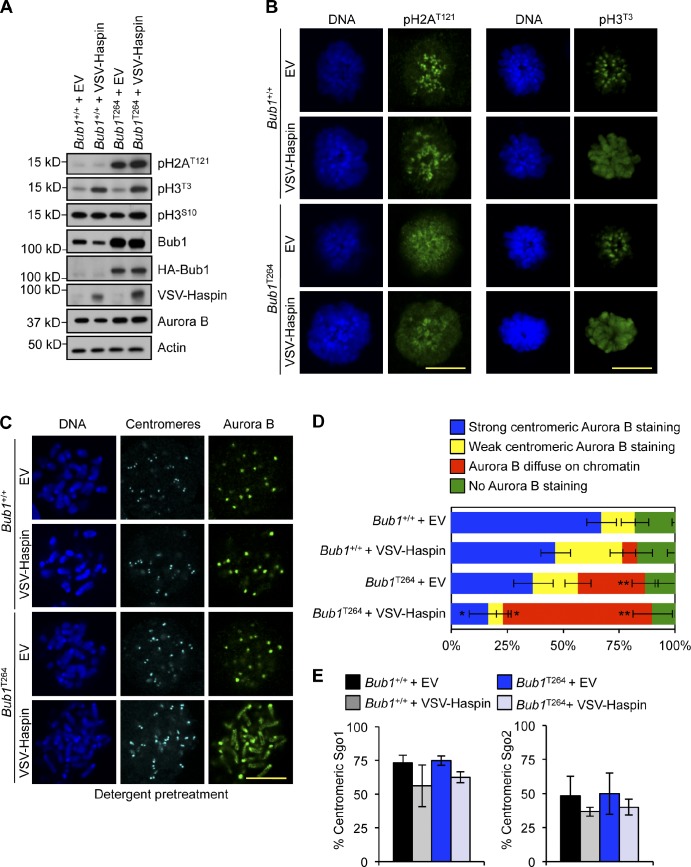
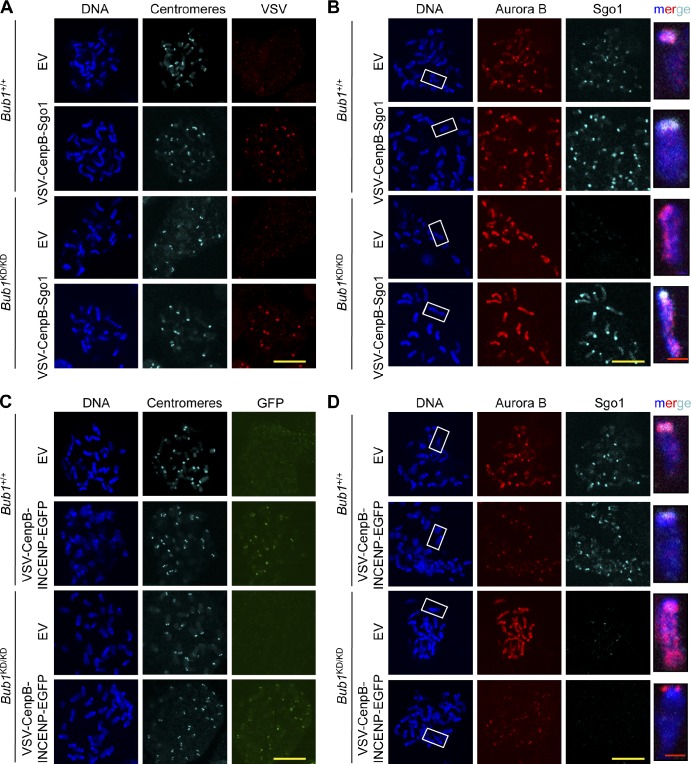
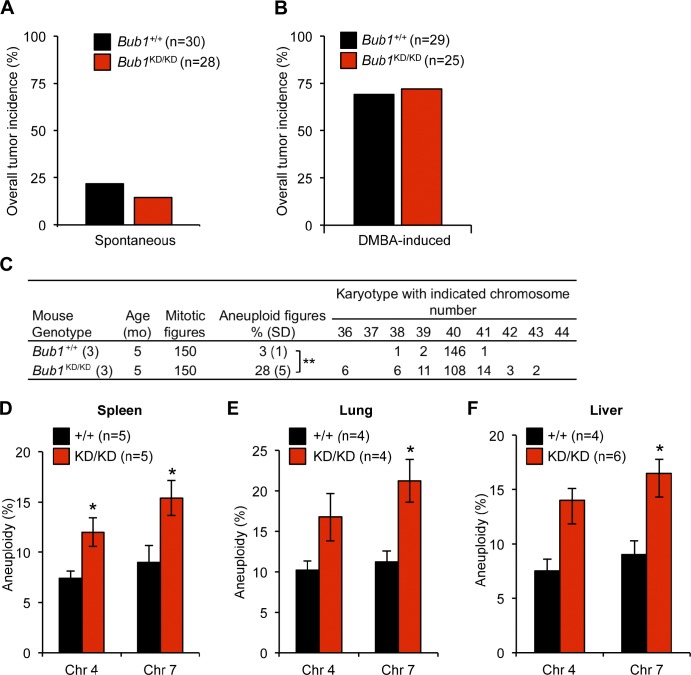
The mitotic checkpoint protein Bub1 is essential for embryogenesis and survival of proliferating cells, and bidirectional deviations from its normal level of expression cause chromosome missegregation, aneuploidy, and cancer predisposition in mice. To provide insight into the physiological significance of this critical mitotic regulator at a modular level, we generated Bub1 mutant mice that lack kinase activity using a knockin gene-targeting approach that preserves normal protein abundance. In this paper, we uncover that Bub1 kinase activity integrates attachment error correction and mitotic checkpoint signaling by controlling the localization and activity of Aurora B kinase through phosphorylation of histone H2A at threonine 121. Strikingly, despite substantial chromosome segregation errors and aneuploidization, mice deficient for Bub1 kinase activity do not exhibit increased susceptibility to spontaneous or carcinogen-induced tumorigenesis. These findings provide a unique example of a modular mitotic activity orchestrating two distinct networks that safeguard against whole chromosome instability and reveal the differential importance of distinct aneuploidy-causing Bub1 defects in tumor suppression.
Figures
References
-
- Bolanos-Garcia V.M., Kiyomitsu T., D’Arcy S., Chirgadze D.Y., Grossmann J.G., Matak-Vinkovic D., Venkitaraman A.R., Yanagida M., Robinson C.V., Blundell T.L. 2009. The crystal structure of the N-terminal region of BUB1 provides insight into the mechanism of BUB1 recruitment to kinetochores. Structure. 17:105–116 10.1016/j.str.2008.10.015 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
