

Central role for endothelial human deneddylase-1/SENP8 in fine-tuning the vascular inflammatory response
- PMID: 23209320
- PMCID: PMC3529810
- DOI: 10.4049/jimmunol.1202041
Central role for endothelial human deneddylase-1/SENP8 in fine-tuning the vascular inflammatory response
Abstract
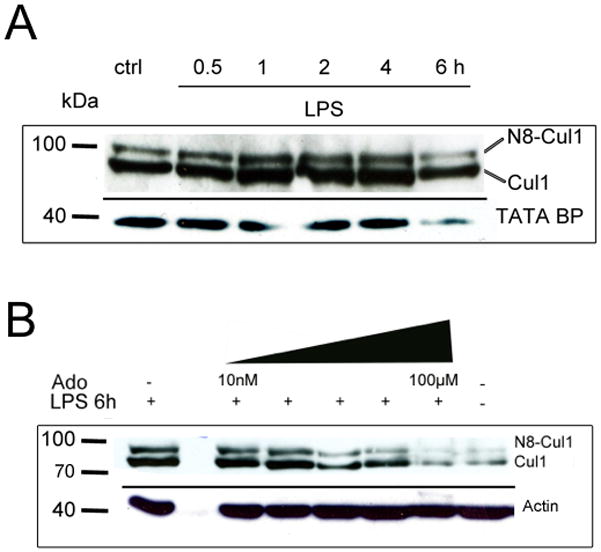
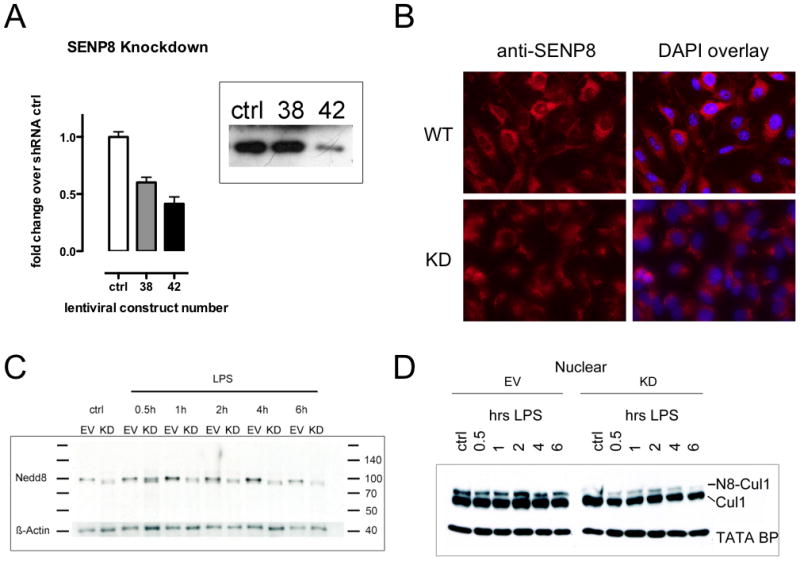
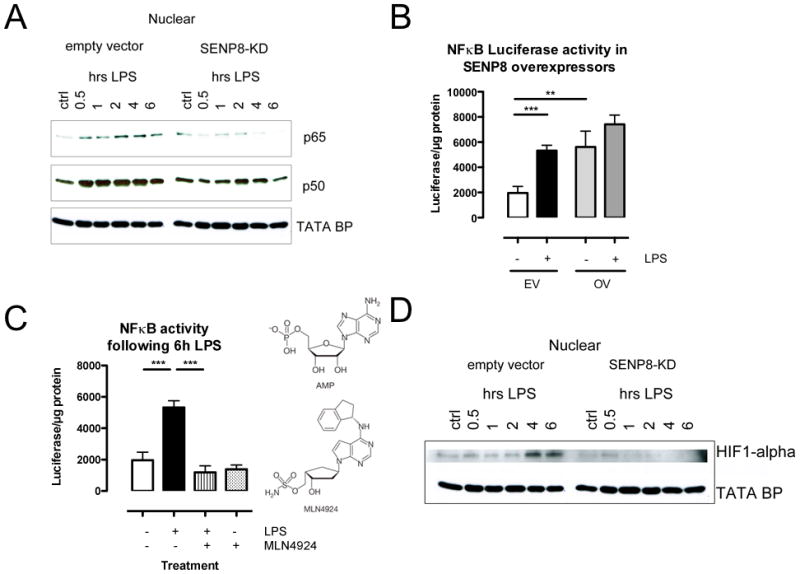
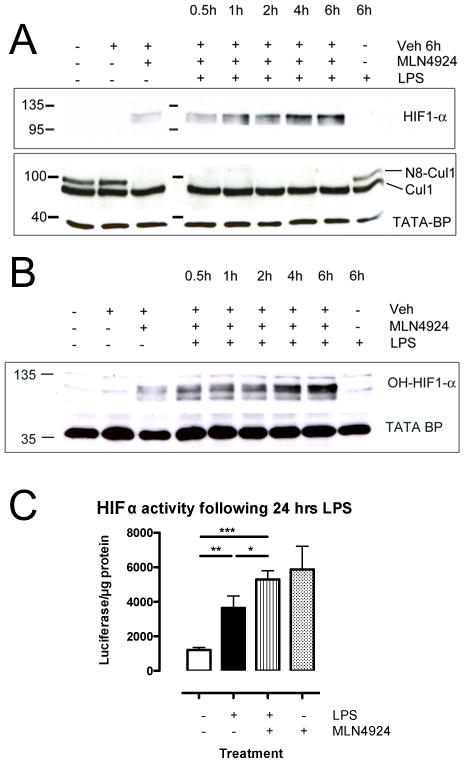
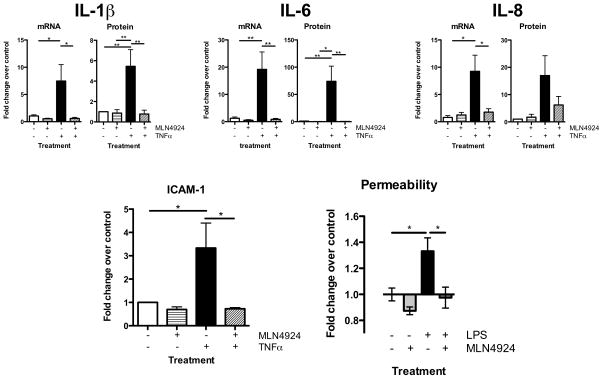
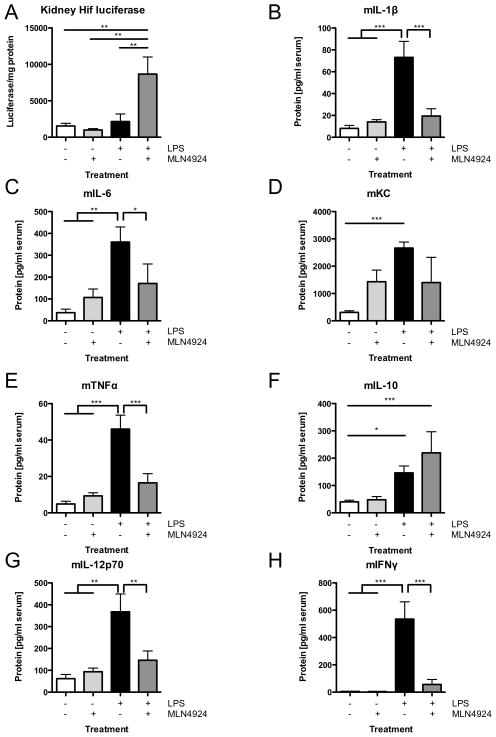
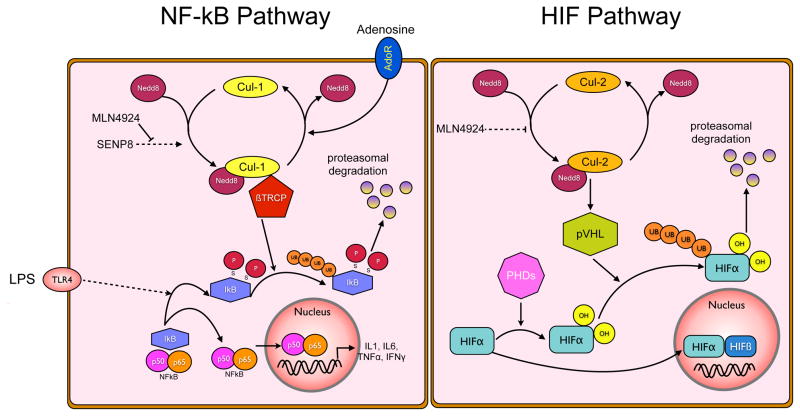
A deeper understanding of the mechanisms that control responses to inflammation is critical to the development of effective therapies. We sought to define the most proximal regulators of the Cullin (Cul)-RING ligases, which play a central role in the stabilization of NF-κB and hypoxia-inducible factor (HIF). In these studies, we identify the human deneddylase-1 (SENP8) as a key regulator of Cul neddylation response in vitro and in vivo. Using human microvascular endothelial cells (HMECs), we examined inflammatory responses to LPS or TNF-α by assessing Cul neddylation status, NF-κB and HIF-1α stabilization, and inflammatory cytokine secretion. HMECs with an intact neddylation pathway showed a time-dependent induction of Cul-1 neddylation, nuclear translocation of NF-κB, stabilization of HIF-1α, and increased NF-κB/HIF-α promoter activity in response to LPS. HMECs lacking SENP8 were unable to neddylate Cul-1 and subsequently were unable to activate NF-κB or HIF-1α. Pharmacological targeting of neddylation (MLN4924) significantly abrogated NF-κB responses, induced HIF-1α promoter activity, and reduced secretion of TNF-α-elicited proinflammatory cytokines. MLN4924 stabilized HIF and abrogated proinflammatory responses while maintaining anti-inflammatory IL-10 responses in vivo following LPS administration. These studies identify SENP8 as a proximal regulator of Cul neddylation and provide an important role for SENP8 in fine-tuning the inflammatory response. Moreover, our findings provide feasibility for therapeutic targeting of the Culs during inflammation.
Conflict of interest statement
Figures
References
-
- Angus DC, Linde-Zwirble WT, Lidicker J, Clermont G, Carcillo J, Pinsky MR. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit Care Med. 2001;29:1303–1310. - PubMed
-
- Martin GS, Mannino DM, Eaton S, Moss M. The epidemiology of sepsis in the United States from 1979 through 2000. N Engl J Med. 2003;348:1546–1554. - PubMed
-
- Paulus P, Jennewein C, Zacharowski K. Biomarkers of endothelial dysfunction: can they help us deciphering systemic inflammation and sepsis? Biomarkers. 2011;16(Suppl 1):S11–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
