

Assessment of human multi-potent hematopoietic stem/progenitor cell potential using a single in vitro screening system
- PMID: 23209758
- PMCID: PMC3509091
- DOI: 10.1371/journal.pone.0050495
Assessment of human multi-potent hematopoietic stem/progenitor cell potential using a single in vitro screening system
Abstract
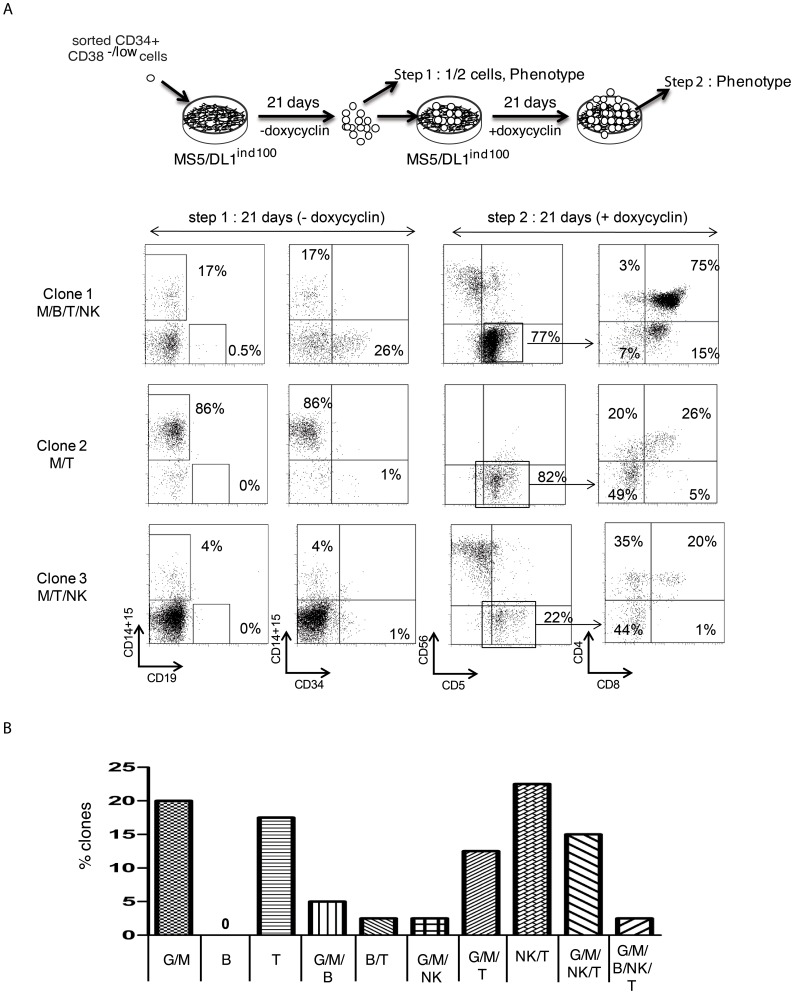
Hematopoietic stem cells are responsible for the generation of the entire blood system through life. This characteristic relies on their ability to self renew and on their multi-potentiality. Thus quantification of the number of hematopoietic stem cells in a given cell population requires to show both properties in the studied cell populations. Although xenografts models that support human hematopoietic stem cells have been described, such in vivo experimental systems remain restrictive for high throughput screening purposes for example. In this work we developed a conditional tetracycline inducible system controlling the expression of the human NOTCH ligand Delta-like 1 in the murine stromal MS5 cells. We cultured hematopoietic immature cells enriched in progenitor/stem cells in contact with MS5 cells that conditionally express Delta-like 1, in conditions designed to generate multipotential lineage differentiation. We show that upon induction or repression of DL1 expression during co-culture, human immature CD34(+)CD38(-/low)(CD45RA(-)CD90(+)) cells can express their B, T, NK, granulo/monocytic and erythroid potentials in a single well, and at the single cell level. We also document the interference of low NOTCH activation with human B and myelo/erythroid lymphoid differentiation. This system represents a novel tool to precisely quantify human hematopoietic immature cells with both lymphoid and myeloid potentials.
Conflict of interest statement
Figures
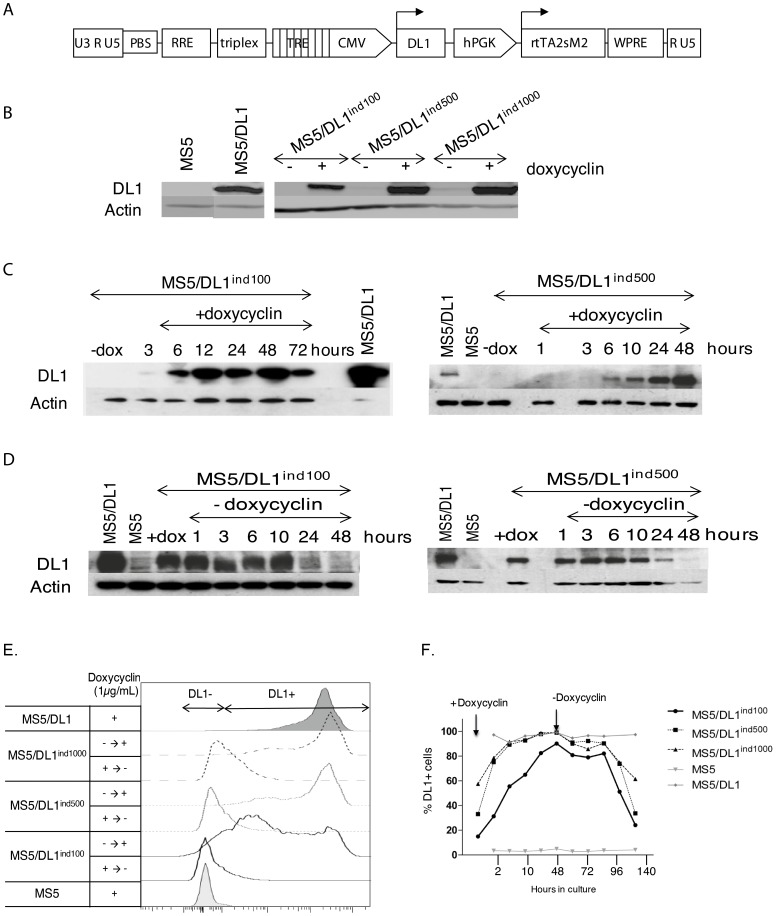
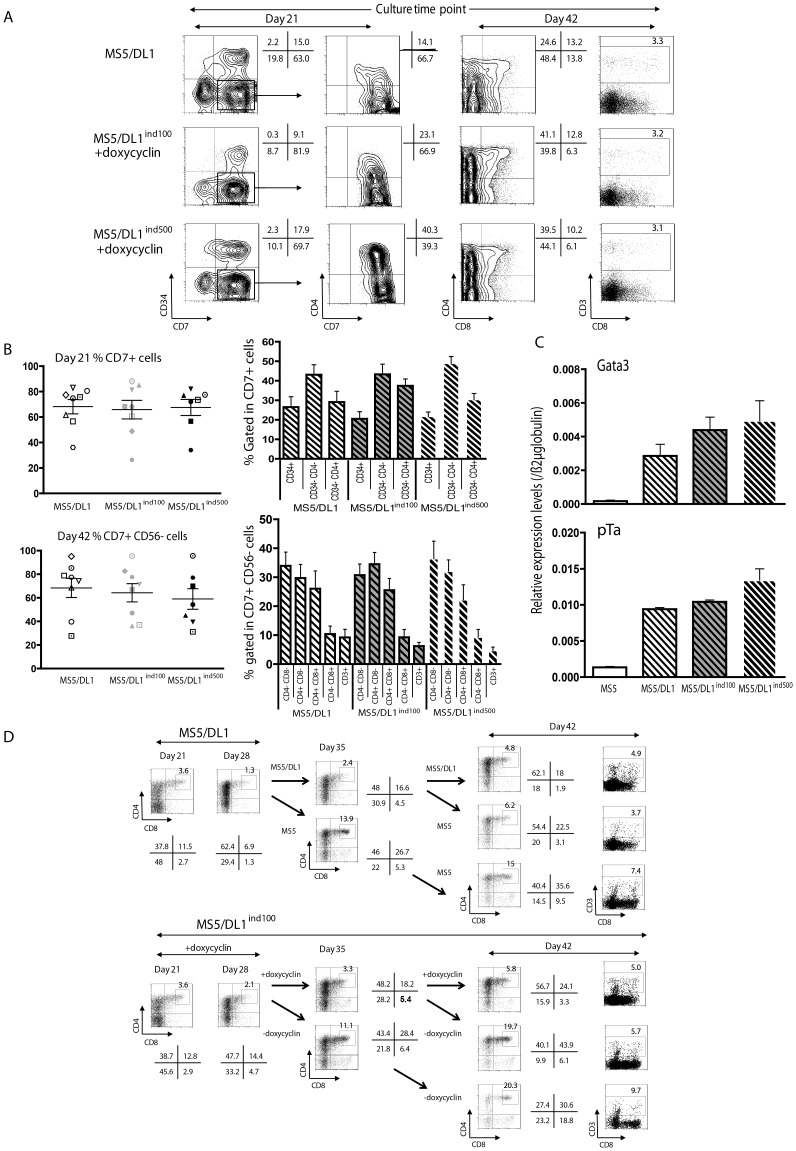
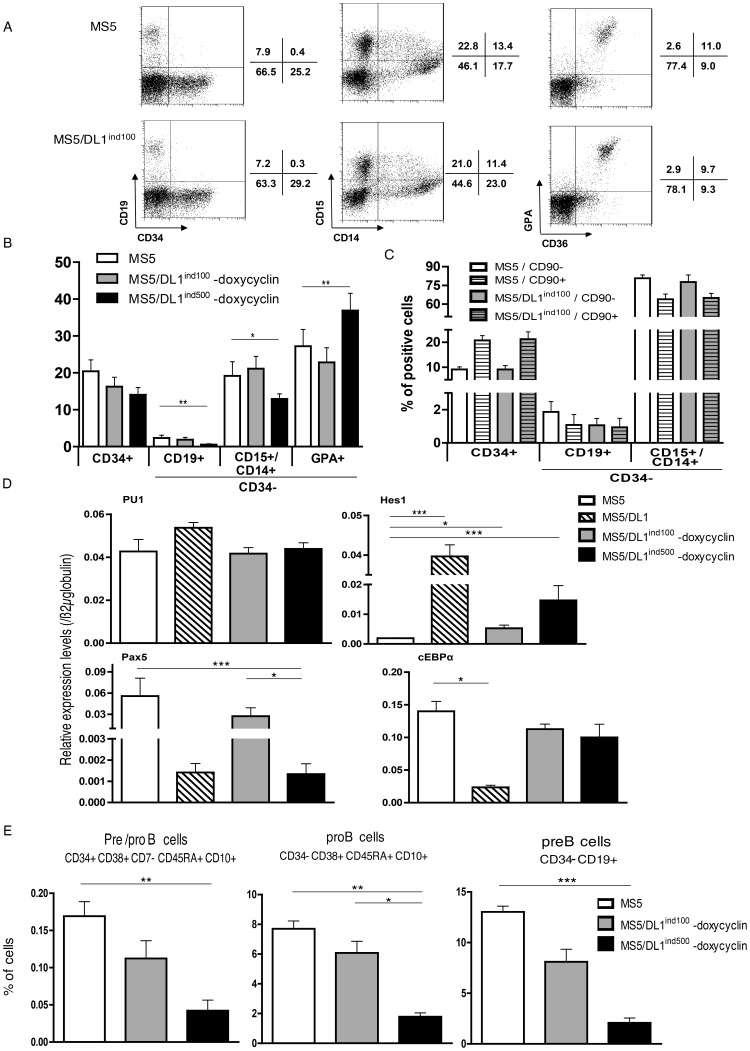
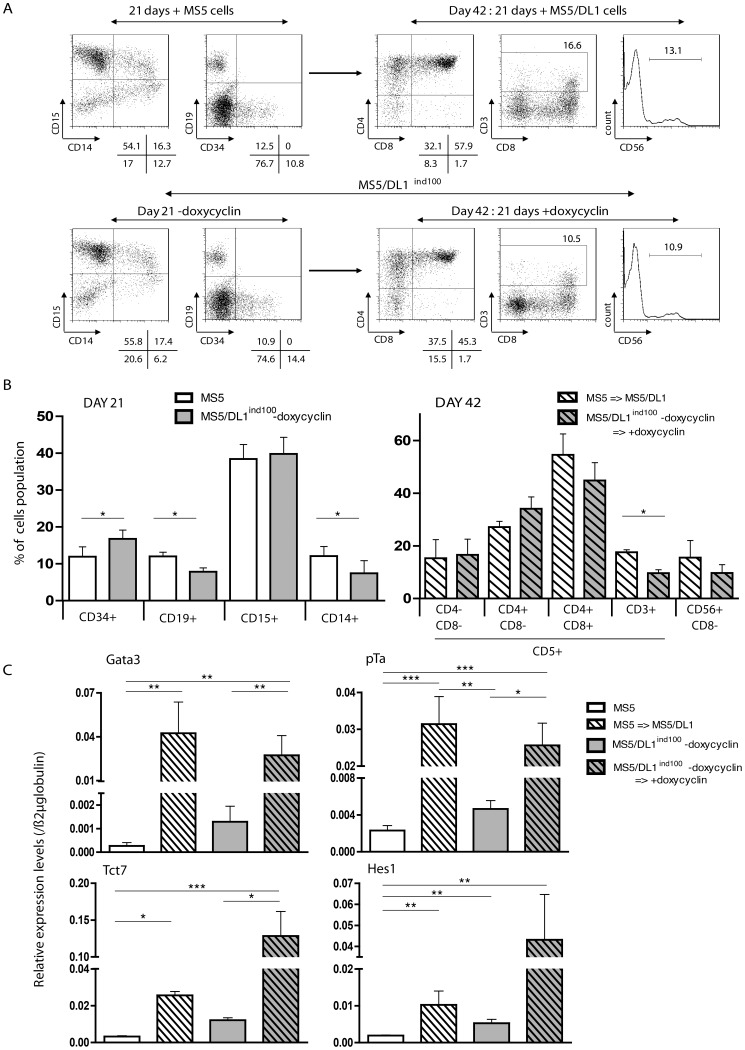
References
-
- Boisset JC, Robin C (2012) On the origin of hematopoietic stem cells: Progress and controversy. Stem Cell Res 8: 1–13. - PubMed
-
- Shultz LD, Ishikawa F, Greiner DL (2007) Humanized mice in translational biomedical research. Nat Rev Immunol 7: 118–130. - PubMed
-
- Coulombel L (2004) Identification of hematopoietic stem/progenitor cells: strength and drawbacks of functional assays. Oncogene 23: 7210–7222. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
