

Dynamic blastomere behaviour reflects human embryo ploidy by the four-cell stage
- PMID: 23212380
- PMCID: PMC3535341
- DOI: 10.1038/ncomms2249
Dynamic blastomere behaviour reflects human embryo ploidy by the four-cell stage
Abstract
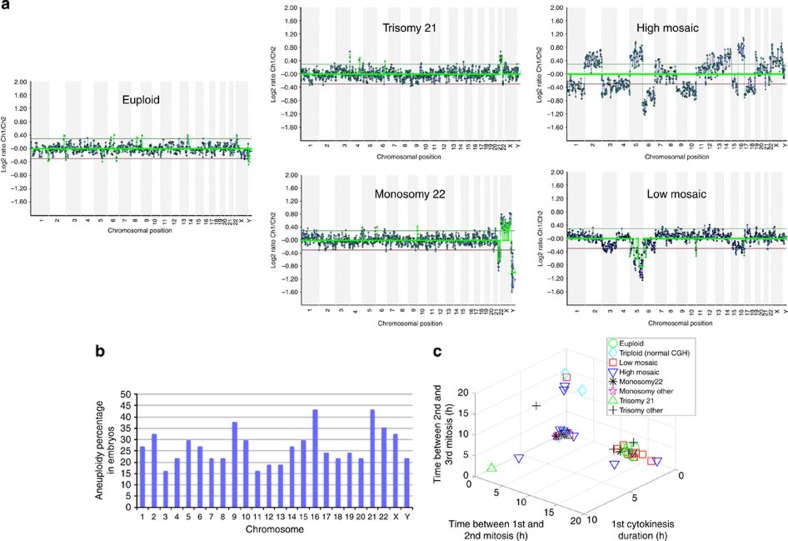
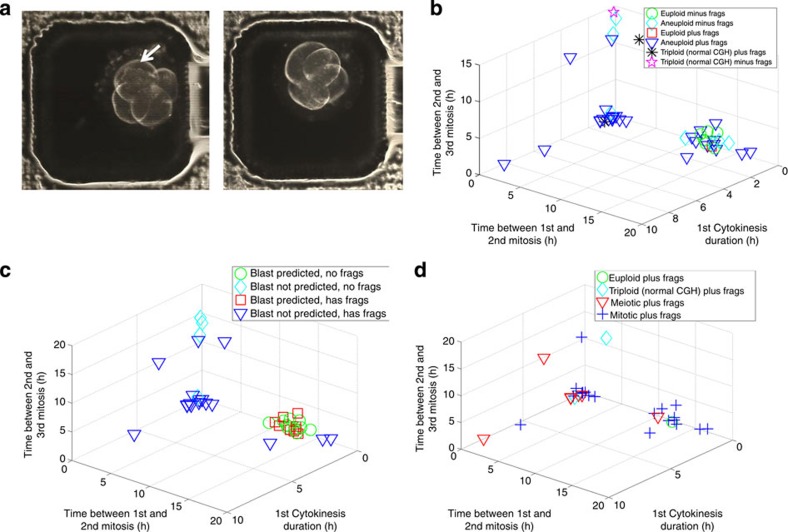
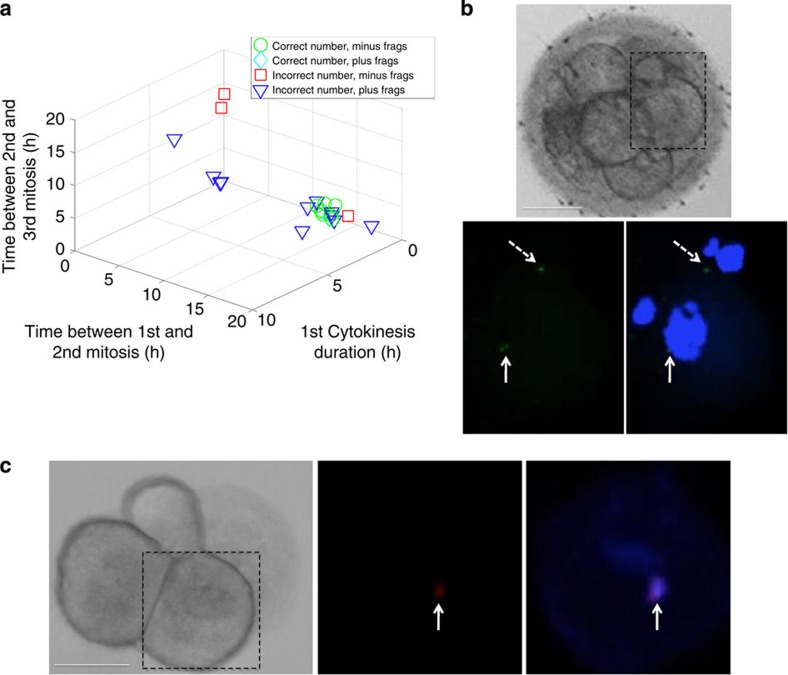
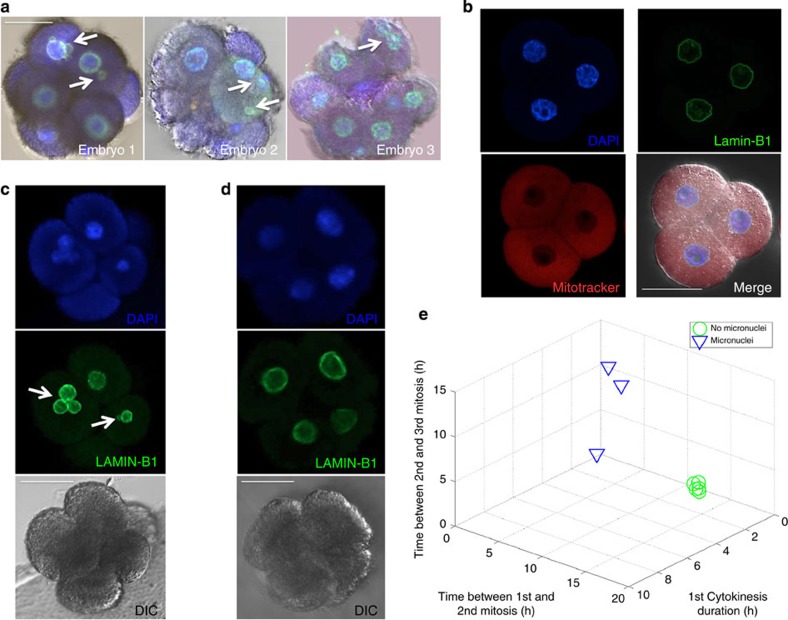
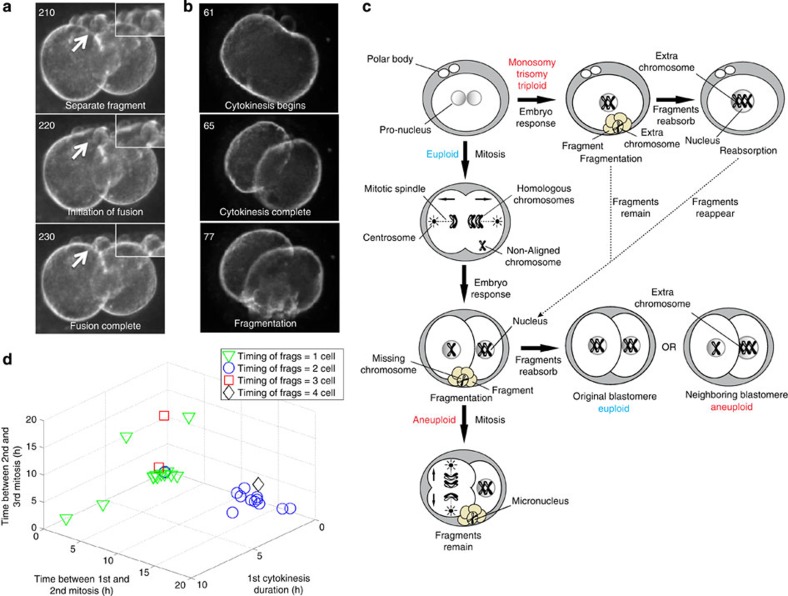
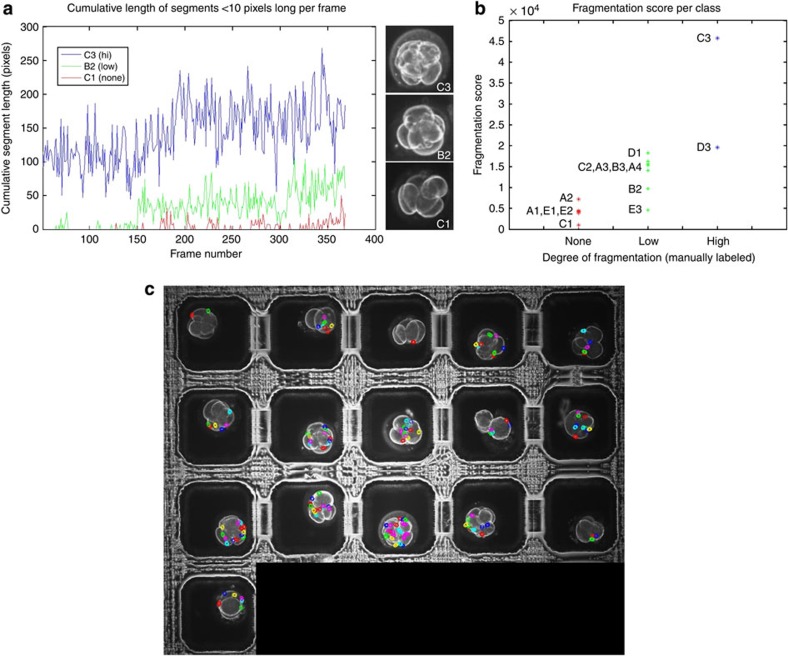
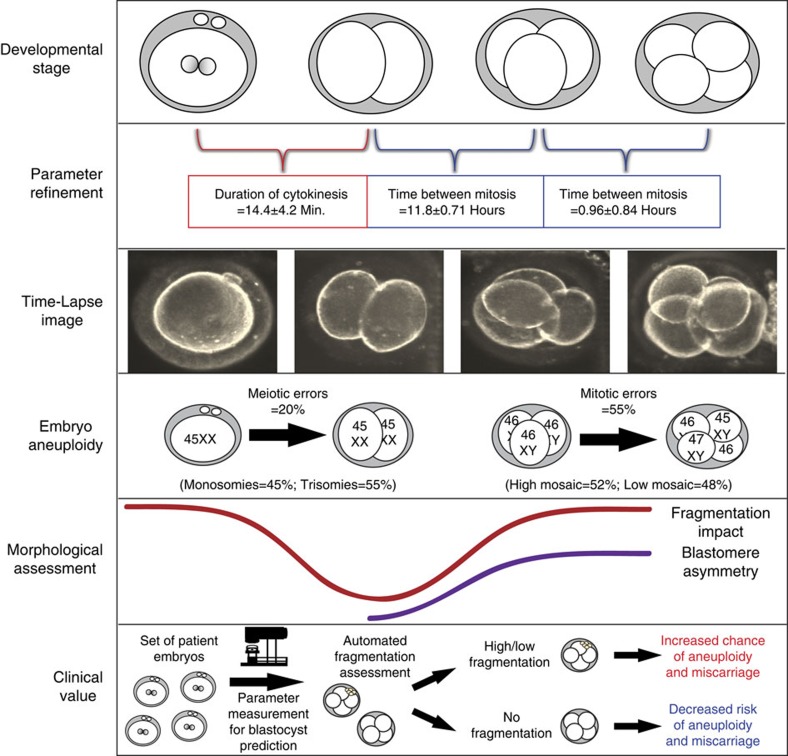
Previous studies have demonstrated that aneuploidy in human embryos is surprisingly frequent with 50-80% of cleavage-stage human embryos carrying an abnormal chromosome number. Here we combine non-invasive time-lapse imaging with karyotypic reconstruction of all blastomeres in four-cell human embryos to address the hypothesis that blastomere behaviour may reflect ploidy during the first two cleavage divisions. We demonstrate that precise cell cycle parameter timing is observed in all euploid embryos to the four-cell stage, whereas only 30% of aneuploid embryos exhibit parameter values within normal timing windows. Further, we observe that the generation of human embryonic aneuploidy is complex with contribution from chromosome-containing fragments/micronuclei that frequently emerge and may persist or become reabsorbed during interphase. These findings suggest that cell cycle and fragmentation parameters of individual blastomeres are diagnostic of ploidy, amenable to automated tracking algorithms, and likely of clinical relevance in reducing transfer of embryos prone to miscarriage.
Conflict of interest statement
This research project was conducted at Stanford University. K.E.L. and F.M. are employees of Auxogyn, Inc. K.E.L., B.B. and R.A.R.P own stock in Auxogyn.
Figures
References
-
- Wong C. et al. Non-invasive imaging of human embryos before embryonic genome activation predicts development to the blastocyst stage. Nat. Biotechnol. 28, 1115–1121 (2010). - PubMed
-
- Dobson A. T. et al. The unique transcriptome through day 3 of human preimplantation development. Hum. Mol. Genet. 13, 1461–1470 (2004). - PubMed
-
- Vanneste E. et al. Chromosome instability is common in human cleavage-stage embryos. Nat. Med. 15, 577–583 (2009). - PubMed
Publication types
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
