

Mitochondrial protein synthesis, import, and assembly
- PMID: 23212899
- PMCID: PMC3512135
- DOI: 10.1534/genetics.112.141267
Mitochondrial protein synthesis, import, and assembly
Abstract
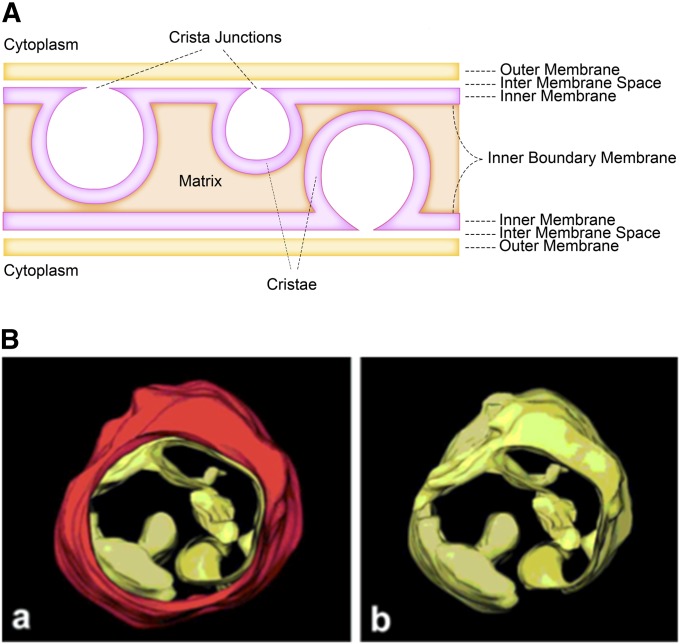
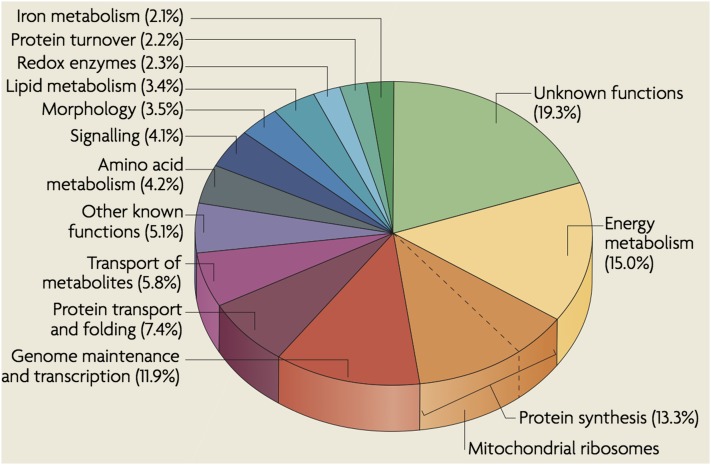
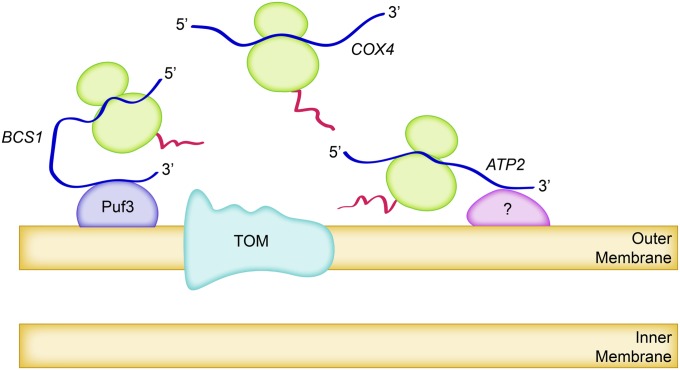
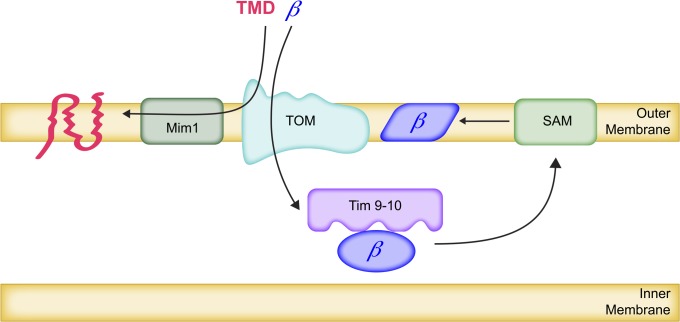
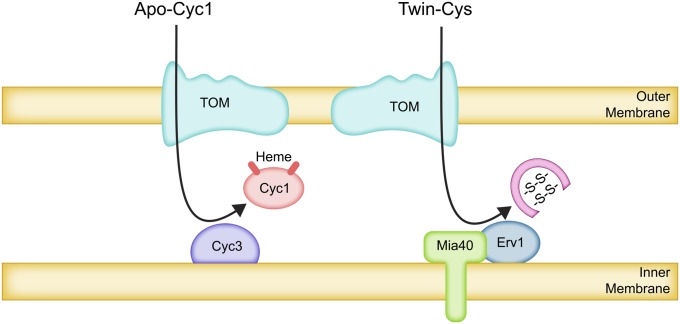
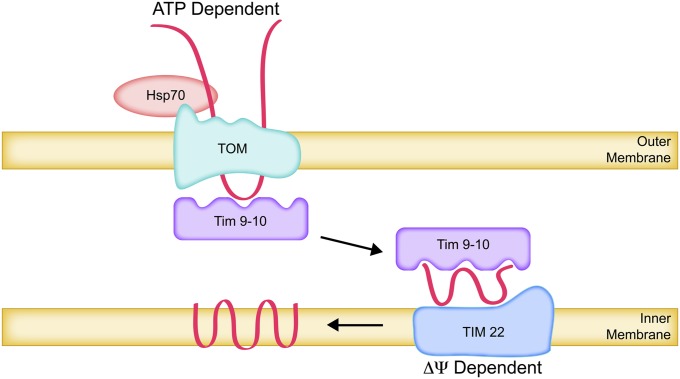
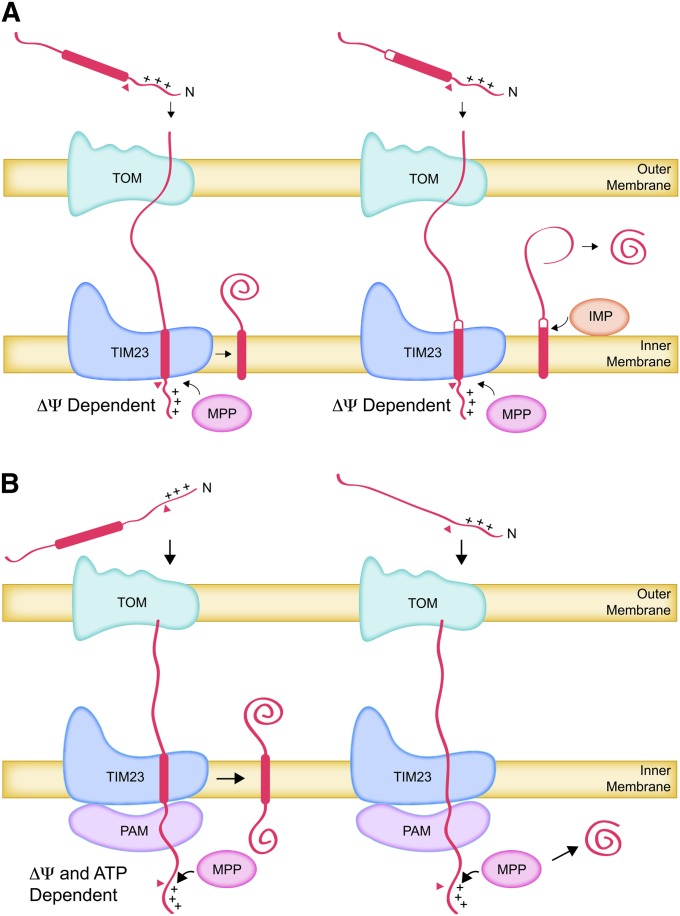
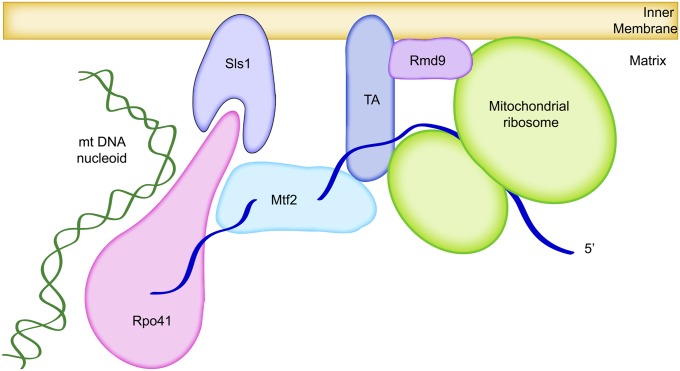
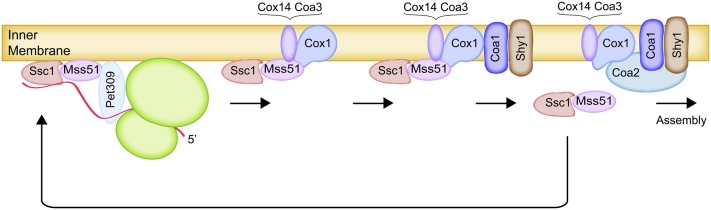
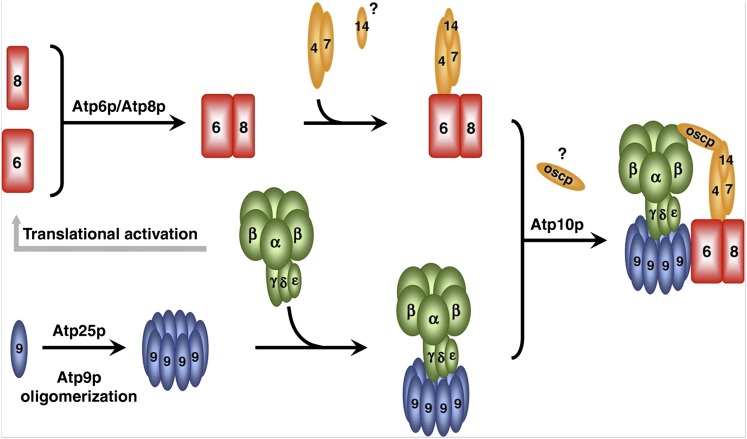
The mitochondrion is arguably the most complex organelle in the budding yeast cell cytoplasm. It is essential for viability as well as respiratory growth. Its innermost aqueous compartment, the matrix, is bounded by the highly structured inner membrane, which in turn is bounded by the intermembrane space and the outer membrane. Approximately 1000 proteins are present in these organelles, of which eight major constituents are coded and synthesized in the matrix. The import of mitochondrial proteins synthesized in the cytoplasm, and their direction to the correct soluble compartments, correct membranes, and correct membrane surfaces/topologies, involves multiple pathways and macromolecular machines. The targeting of some, but not all, cytoplasmically synthesized mitochondrial proteins begins with translation of messenger RNAs localized to the organelle. Most proteins then pass through the translocase of the outer membrane to the intermembrane space, where divergent pathways sort them to the outer membrane, inner membrane, and matrix or trap them in the intermembrane space. Roughly 25% of mitochondrial proteins participate in maintenance or expression of the organellar genome at the inner surface of the inner membrane, providing 7 membrane proteins whose synthesis nucleates the assembly of three respiratory complexes.
Figures
References
-
- Abe Y., Shodai T., Muto T., Mihara K., Torii H., et al. , 2000. Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell 100: 551–560 - PubMed
-
- Ackerman S. H., Tzagoloff A., 2005. Function, structure, and biogenesis of mitochondrial ATP synthase. Prog. Nucleic Acid Res. Mol. Biol. 80: 95–133 - PubMed
-
- Ackerman S. H., Gatti D. L., Gellefors P., Douglas M. G., Tzagoloff A., 1991. ATP13, a nuclear gene of Saccharomyces cerevisiae essential for the expression of subunit 9 of the mitochondrial ATPase. FEBS Lett. 278: 234–238 - PubMed
-
- Ades I. Z., Butow R. A., 1980. The products of mitochondria-bound cytoplasmic polysomes in yeast. J. Biol. Chem. 255: 9918–9924 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
