

Mutations of the Ca2+-sensing stromal interaction molecule STIM1 regulate Ca2+ influx by altered oligomerization of STIM1 and by destabilization of the Ca2+ channel Orai1
- PMID: 23212906
- PMCID: PMC3548475
- DOI: 10.1074/jbc.M112.417246
Mutations of the Ca2+-sensing stromal interaction molecule STIM1 regulate Ca2+ influx by altered oligomerization of STIM1 and by destabilization of the Ca2+ channel Orai1
Abstract
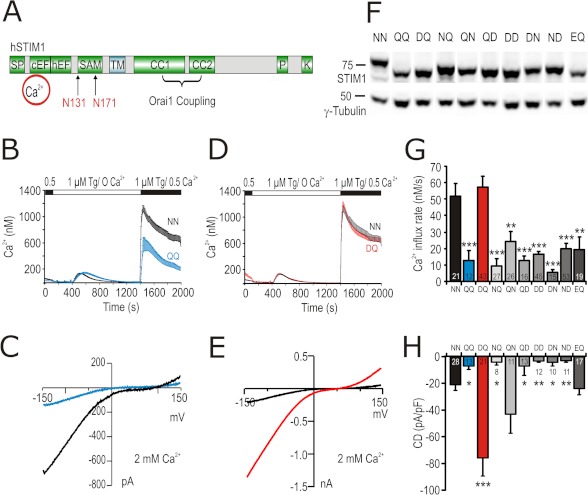
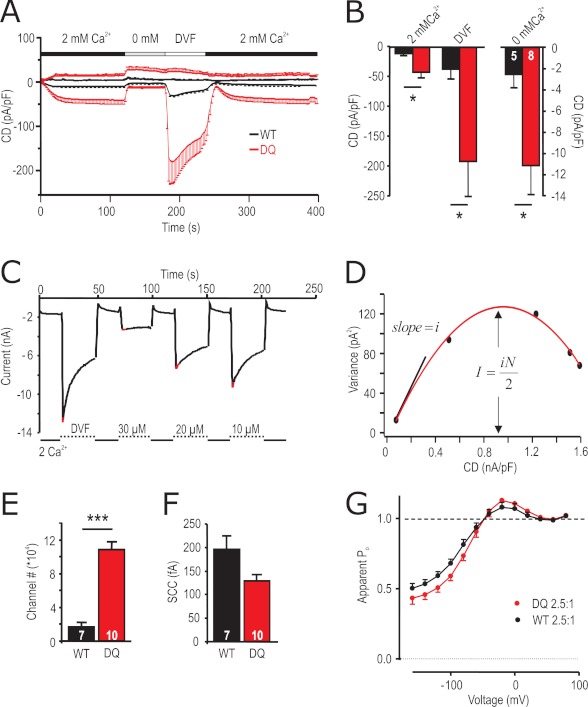
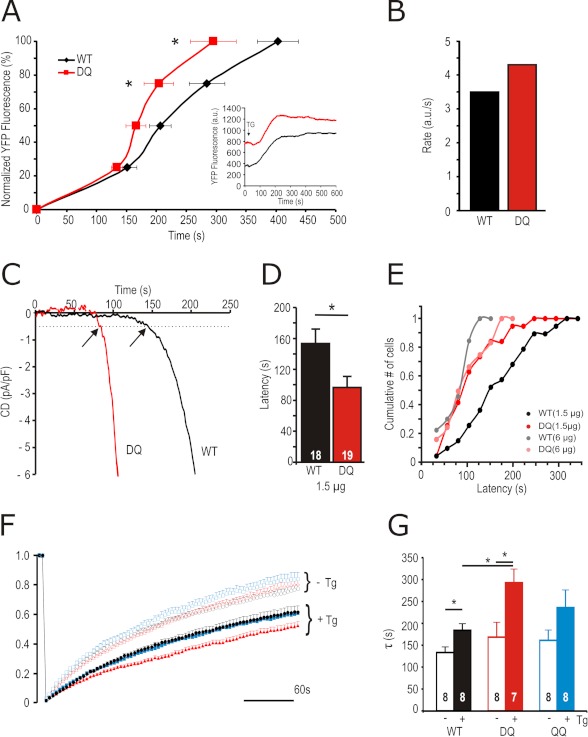
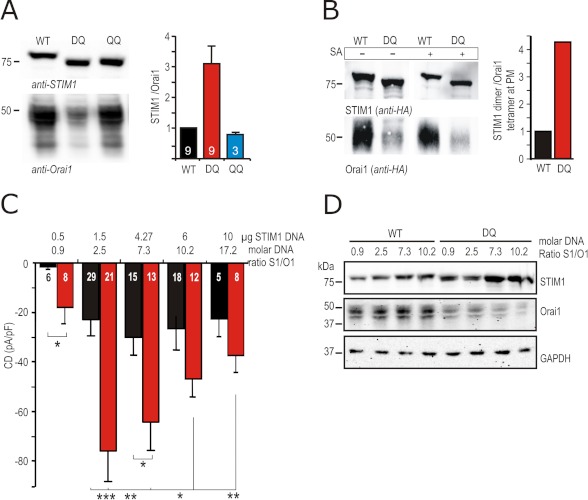
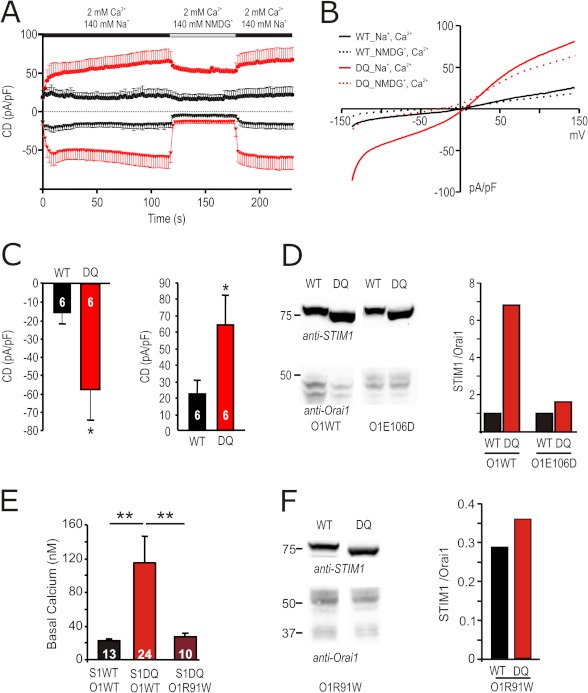
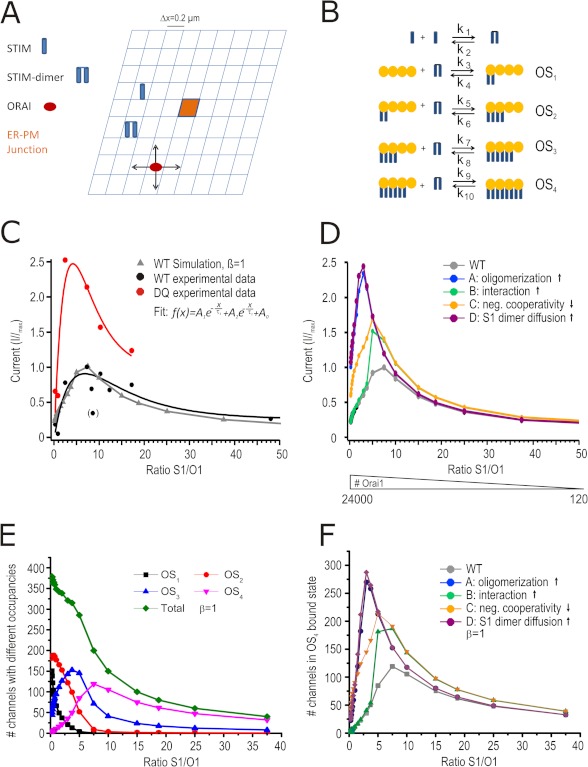
A drop of endoplasmic reticulum Ca(2+) concentration triggers its Ca(2+) ssensor protein stromal interaction molecule 1 (STIM1) to oligomerize and accumulate within endoplasmic reticulum-plasma membrane junctions where it activates Orai1 channels, providing store-operated Ca(2+) entry. To elucidate the functional significance of N-glycosylation sites of STIM1, we created different mutations of asparagine-131 and asparagine-171. STIM1 NN/DQ resulted in a strong gain of function. Patch clamp, Total Internal Reflection Fluorescent (TIRF) microscopy, and fluorescence recovery after photobleaching (FRAP) analyses revealed that expression of STIM1 DQ mutants increases the number of active Orai1 channels and the rate of STIM1 translocation to endoplasmic reticulum-plasma membrane junctions with a decrease in current latency. Surprisingly, co-expression of STIM1 DQ decreased Orai1 protein, altering the STIM1:Orai1 stoichiometry. We describe a novel mathematical tool to delineate the effects of altered STIM1 or Orai1 diffusion parameters from stoichiometrical changes. The mutant uncovers a novel mechanism whereby "superactive" STIM1 DQ leads to altered oligomerization rate constants and to degradation of Orai1 with a change in stoichiometry of activator (STIM1) to effector (Orai1) ratio leading to altered Ca(2+) homeostasis.
Figures
References
-
- Maul-Pavicic A., Chiang S. C., Rensing-Ehl A., Jessen B., Fauriat C., Wood S. M., Sjöqvist S., Hufnagel M., Schulze I., Bass T., Schamel W. W., Fuchs S., Pircher H., McCarl C. A., Mikoshiba K., Schwarz K., Feske S., Bryceson Y. T., Ehl S. (2011) ORAI1-mediated calcium influx is required for human cytotoxic lymphocyte degranulation and target cell lysis. Proc. Natl. Acad. Sci. U.S.A. 108, 3324–3329 - PMC - PubMed
-
- Qu B., Al-Ansary D., Kummerow C., Hoth M., Schwarz E. C. (2011) ORAI-mediated calcium influx in T cell proliferation, apoptosis and tolerance. Cell Calcium 50, 261–269 - PubMed
-
- Feske S., Gwack Y., Prakriya M., Srikanth S., Puppel S. H., Tanasa B., Hogan P. G., Lewis R. S., Daly M., Rao A. (2006) A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 441, 179–185 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
